


All Cellular Microbiology Articles
Export Citations
Download PDFs
-
Issue Informationfree
-
Cover Image: Entry of the Varicellovirus Canid herpesvirus 1 into Madin–Darby canine kidney epithelial cells is pH-independent and occurs via a macropinocytosis-like mechanism but without increase in fluid uptake (Cellular Microbiology 12/2021)free
Graphical Abstract
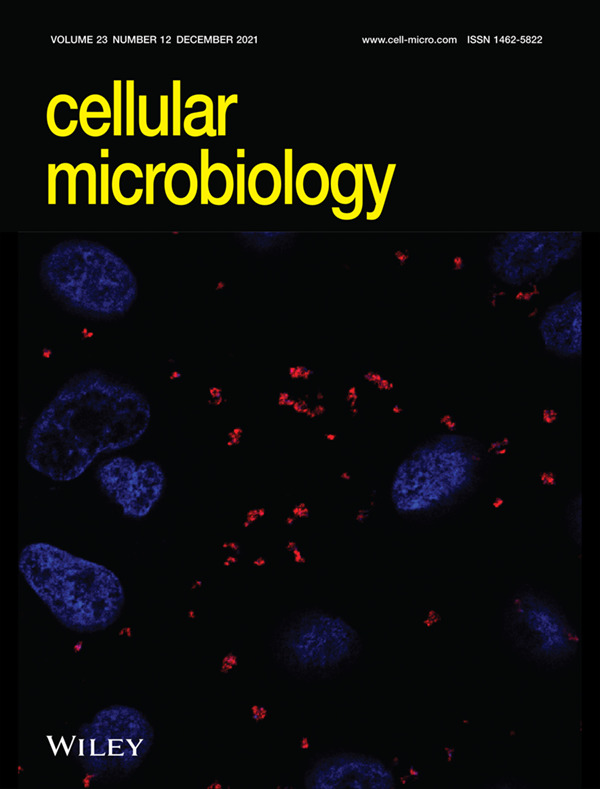
Confocal micrograph showing Canid herpesvirus 1 (red) bound to MDCK cells, and DAPI-stained nuclei (blue). Primary amines of viral glycoproteins were labelled with Alexa Fluor succinimidyl esters 568 dye. For further details, readers are referred to the article by Eisa et al. on p. e13398 of this issue.
-
Vam6/Vps39/TRAP1-domain proteins influence vacuolar morphology, iron acquisition and virulence in Cryptococcus neoformansfull
Graphical Abstract
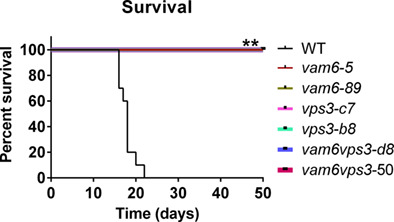
The pathogenic fungus Cryptococcus neoformans must overcome iron limitation to cause disease in mammalian hosts. Two Vam6/Vps39/TRAP1 domain-containing proteins, Vps3 and Vam6, are required for robust growth on haem as well as proper regulation of iron homeostasis. Two independent deletion mutants for each gene, or double mutants lacking both genes, are unable to cause disease in a mouse inhalation model of cryptococcosis.
-
Hepatitis B virus envelope proteins can serve as therapeutic targets embedded in the host cell plasma membraneoa
Graphical Abstract
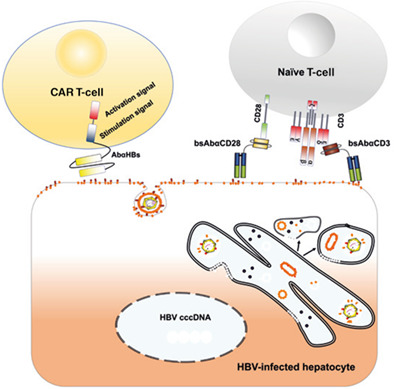
Surface immunoprecipitation of HBS confirmed that membrane-associated HBs remains correctly folded in HBV-replicating cells in cell culture. MoMab coated onto superparamagnetic iron oxide nanoparticles allowed to detect membrane-associated HBs after HBV infection by electron microscopy in distinct stretched of the hepatocyte plasma membrane. Last but not least, we demonstrate that HBs located on the cell surface allow therapeutic targeting of HBV-positive cells by T-cells either engrafted with a chimeric antigen receptor or redirected by bispecific, T-cell engager antibodies.
-
Chlamydia and HPV induce centrosome amplification in the host cell through additive mechanismsfull
Graphical Abstract
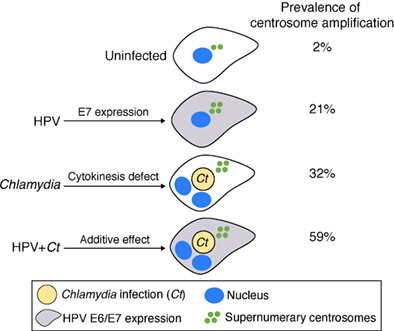
HPV and Chlamydia have additive effects on the prevalence of centrosome amplification when present in the same host cell. While HPV primarily causes centriole overduplication through expression of the oncoprotein E7, Chlamydia causes cytokinesis defects, which then lead to centrosome amplification and multinucleation in the host cell.
-
Entry of the Varicellovirus Canid herpesvirus 1 into Madin–Darby canine kidney epithelial cells is pH-independent and occurs via a macropinocytosis-like mechanism but without increase in fluid uptakefull
Graphical Abstract
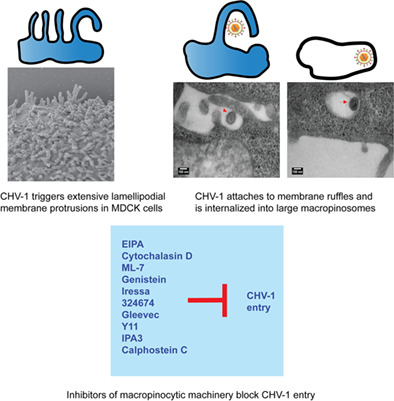
The entry pathway of Canid herpesvirus 1 into MDCK cells was shown to be via a macropinocytosis-like mechanism. CHV-1 induces membrane ruffles and internalises within large uncoated endocytic vacuoles. Pharmacological inhibitors targeting the macropinocytic machinery blocked virus entry, while in contrast, inhibiting endosomal acidification had no effect on the ability of the virus to infect MDCK cells.
-
Issue Informationfree
-
Cover Image: The fungivorous amoeba Protostelium aurantium targets redox homeostasis and cell wall integrity during intracellular killing of Candida parapsilosis (Cellular Microbiology 11/2021)free
Graphical Abstract
The fungivorous amoeba Protostelium aurantium feeds on a wide range of fungal species. The image shows amoebae digesting GFP-expressing cells of the human pathogenic yeast Candida parapsilosis. For further details, readers are referred to the article by Radosa et al. on p. e13389 of this issue.
-
Dengue virus replication enhances labile zinc pools by modulation of ZIP8oa
Graphical Abstract
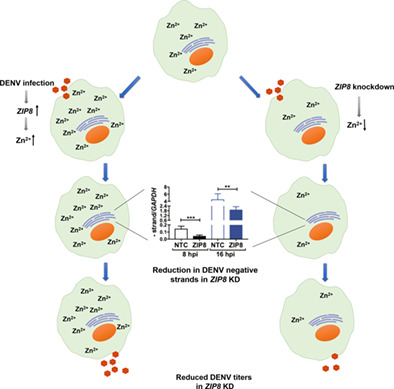
Dengue virus infection was shown to induce intracellular labile zinc pools which colocalized with ER and Golgi markers. ZIP8 levels were enhanced during early stages of dengue virus replication and silencing ZIP8 blocked increase in labile zinc levels and inhibited viral replication.
-
Issue Informationfree
-
Cover Image: Candidalysin delivery to the invasion pocket is critical for host epithelial damage induced by Candida albicans (Cellular Microbiology 10/2021)free
Graphical Abstract
Candida albicans hyphae secreting the peptide toxin candidalysin (green) during invasion of epithelial cells. The toxin accumulates in the “invasion pocket” and damages the host cell. For further details, readers are referred to the article by Mogavero et al. on p. e13378 of this issue.
-
Egress of archaeal virusesfree
Graphical Abstract
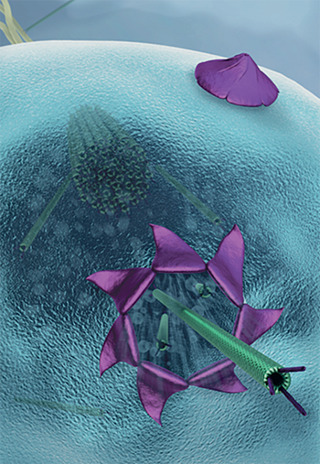
Archaeal viruses infecting Crenarchaeota exploit a unique egress mechanism based on the formation and opening of large pyramidal portals on the host cell surface.
-
Blockade of endoplasmic reticulum stress-induced cell death by Ureaplasma parvum vacuolating factorfull
Graphical Abstract
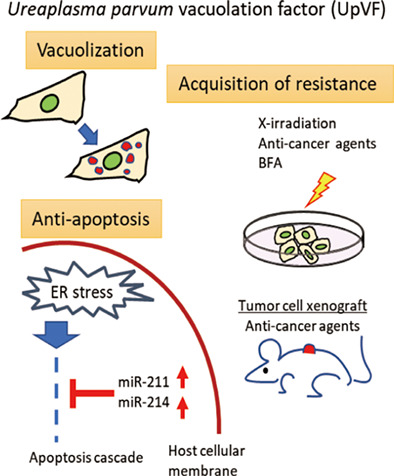
A novel virulence factor, Ureaplasma parvum vacuolating factor (UpVF), was identified. UpVF induced cell vacuolation and triggered ER stress in host cells. In contrast, UpVF blocked the downstream apoptotic cascade through the upregulation of host cellular miR-211 and miR-214. UpVF expressing tumour cell lines showed resistance to anticancer drugs and X-ray irradiation. Xenograft of UpVF expressing tumour cells reduced the efficacy of anticancer agents in nude mice.
-
BSC2 induces multidrug resistance via contributing to the formation of biofilm in Saccharomyces cerevisiaefull
Graphical Abstract
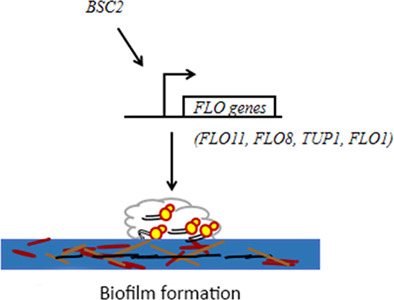
BSC2 promotes biofilm formation in a FLO pathway-dependent manner to induce MDR.
-
Rickettsia conorii survival in THP-1 macrophages involves host lipid droplet alterations and active rickettsial protein productionfull
Graphical Abstract
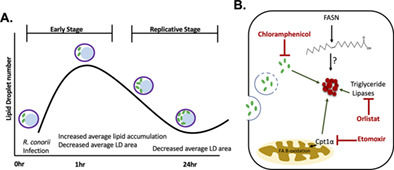
Rickettsia conorii infection triggers dynamic changes to host cell lipid content, which can be visualised by an increase in lipid droplet (LD) production early in infection and a subsequent decrease in average LD area throughout infection (a). Pharmacological inhibition of lipid catabolic pathways perturbs these infection-induced alterations in LDs, suggesting that fatty acid oxidation (FAO) and triglyceride lipases are involved in stimulating the LD modulation seen at early stages. These alterations in LDs observed early in the infection process are driven by R. conorii de novo protein synthesis (b).
-
The fungivorous amoeba Protostelium aurantium targets redox homeostasis and cell wall integrity during intracellular killing of Candida parapsilosisfull
Graphical Abstract
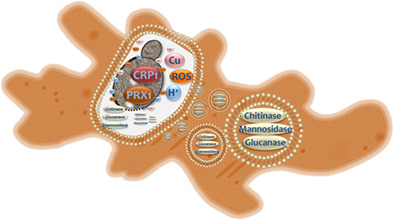
The amoeba Protostelium aurantium feeds on fungi, including human pathogenic species such as Candida parapsilosis. Following ingestion, yeast cells are exposed to reactive oxygen species (ROS) in an acidified phagosome (H+). A copper exporter (CRP1) and a peroxiredoxin (PRX1) contribute to defence while lysis occurs via hydrolytic enzymes from intracellular vesicles of the amoeba.
-
Porphyromonas gingivalis induces penetration of lipopolysaccharide and peptidoglycan through the gingival epithelium via degradation of coxsackievirus and adenovirus receptorfull
Graphical Abstract
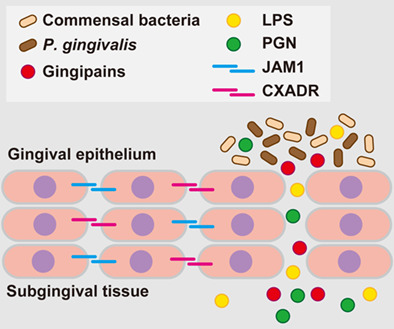
Porphyromonas gingivalis, a major pathogen of human periodontitis, secretes gingipains, degrading coxsackievirus and adenovirus receptor (CXADR) and junctional adhesion molecule 1 (JAM1), gingival epithelial barrier proteins, and increasing permeability to gingipains. Subsequently, gingipains become translocated to deeper epithelium for additional degradation of CXADR and JAM1, thus allowing lipopolysaccharide (LPS) and peptidoglycan (PGN) to penetrate the gingival epithelium and reach subepithelial tissues.
-
Zinc finger proteins of Plasmodium falciparumoa
Graphical Abstract
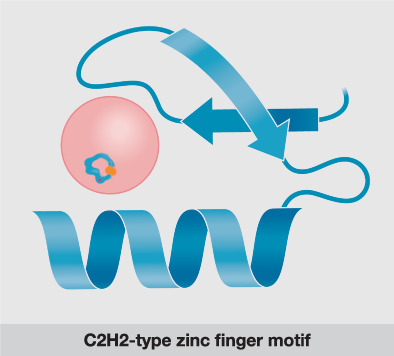
Schematic of a C2H2-type zinc finger motif comprising a two-stranded antiparallel β-sheet and an α-helix coordinated by a zinc ion. The zinc ion is depicted by a Plasmodium falciparum ring stage. In the ring stages, C2H2-type zinc finger proteins are mainly involved in cell cycle regulation and mRNA metabolism.
-
Issue Informationfree