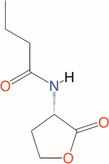
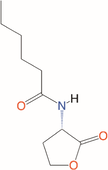
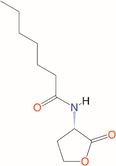
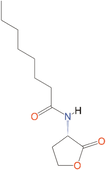
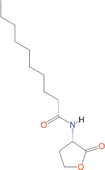
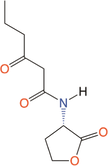
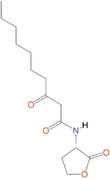
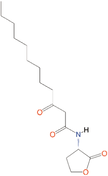
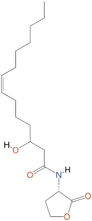
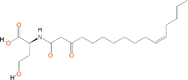
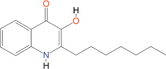
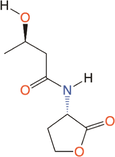
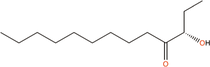
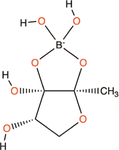
Quorum quenching and quorum sensing inhibitors show significant potential in regulating bacterial quorum sensing systems and have been widely applied across various fields, including cancer treatment, antimicrobial resistance, marine management, microplastic reduction, hydrogel technology, and nanomaterials development.
ABSTRACT
The study of quorum sensing (QS) has gained critical importance, offering insights into bacterial and microorganism communication. QS, regulated by autoinducers, synchronizes collective bacterial behaviors across diverse chemical signals and target genes. This review highlights innovative approaches to regulating QS, emphasizing the potential of quorum quenching and QS inhibitors to mitigate bacterial pathogenicity. These strategies have shown promise in aquaculture and plant resistance, disrupting QS pathways to combat infections. QS also provides opportunities for developing biosensors for early disease detection and preventing biofilm formation, which is critical to overcoming antimicrobial resistance. The applications of QS extend to cancer therapy, with targeted drug delivery systems utilizing QS mechanisms. Advancements in QS regulation, such as the use of nanomaterials, hydrogels, and microplastics, provide novel methods to modulate QS systems. This review explores the latest developments in QS, recognizing its significance in controlling bacterial behavior and its broad impacts on human health and disease management. Integrating these insights into therapeutic strategies and diagnostics represents a pivotal opportunity for medical progress.
1 Introduction
Quorum sensing (QS) is a complex communication system among bacteria. QS enables bacterial groups to synchronously coordinate their behavior in response to fluctuations in population density and species composition in neighboring communities. The production, detection, and response to extracellular signaling molecules known as autoinducers are central to this process (Waters and Bassler 2005). As the bacterial population increases, the concentration of autoinducer molecules rises. Once a critical concentration threshold is attained, the bacteria can detect autoinducer molecules and initiate a specific response. QS is essential for various finely adapted behaviors, such as bacterial virulence (Winson et al. 1995), biofilm formation (Cvitkovitch et al. 2003; Solano et al. 2014; L. Xu et al. 2006), drug resistance (Pumbwe et al. 2008), bioluminescence (Engebrecht et al. 1983; Engebrecht and Silverman 1984, 1987), and response to environmental changes (Lee and Zhang 2015). With the advent of QS systems, bacteria are no longer regarded as single-celled living organisms that can only carry out simple processes. Most processes regulated by QS are ineffective when conducted by a single bacterium acting in isolation. Thus, QS enables bacteria to cooperate among individual cells, thereby achieving high levels of coordination. There are differences between QS systems due to adaptations for specific species in particular living conditions. In diverse bacterial species, autoinducers exhibit high specificity, with the type of signal, outputs, transduction mechanisms, and receptors all reflecting the distinct biology of each microorganism (Whiteley et al. 2018).
In the 1960s and 1970s, the first reports of QS revealed that extracellular molecules were essential for competence in Streptococcus pneumoniae (Tomasz 1965) and bioluminescence in marine pathogenic bacteria (Nealson et al. 1970). In Vibrio fischeri, the production of light is closely related to the cell density (Bassler et al. 1994). Bioluminescence is expressed only when the density of V. fischeri cells reaches a sufficiently high level. In the 1980s, the discovery of the genes luxI and luxR, which encode bioluminescence and the autoinducer 3-oxo-C6-HSL used in this system, respectively, provided a complete picture of the QS system (Eberhard et al. 1981; Engebrecht et al. 1983). By the 1990s, DNA sequencing unveiled LuxI and LuxR homologs in various bacteria, leading to the concept of QS (Fuqua et al. 1994). This communication system, which is present in both Gram-negative and Gram-positive bacteria, was initially studied in pathogenic microbes, after which the research focus shifted to the broader implications of bacterial communication. Further research uncovered the autoinducer-2 (AI-2) in Vibrio harveyi (X. Chen et al. 2002), expanding QS beyond bacterial species to include multispecies and eukaryotes (Federle and Bassler 2003). Interspecies communication makes it possible to artificially intervene in bacterial communication within the human body. Later reports demonstrated that in many animal and plant pathogens, the virulence of QS mutants was greatly reduced (Balaban and Novick 1995). Through employing chemical intervention to inhibit the communication between bacteria (Deng et al. 2020) or between bacteria and viruses (Silpe et al. 2023) in the human body, it is possible to use QS to treat diseases.
2 Gram-Negative Bacteria
Engebrecht et al.'s research on V. fischeri laid the foundation for studying QS in Gram-negative bacteria (Engebrecht et al. 1983; Engebrecht and Silverman 1984, 1987) and identified the LuxI–LuxR system and acyl-homoserine lactone (AHL) signaling molecules. The composition of the acyl chain varies from 4 to 16 carbon atoms (Table 1). The signaling molecules exhibit a high degree of specificity due to their structural features (Watson et al. 2002). The most prevalent signaling molecule found in Gram-negative bacteria is the N-acyl-homoserine lactone derivative (Whitehead et al. 2001). Table 2 lists the diverse and complex QS communication networks of Gram-negative bacteria.
Table 1.
List the abbreviations and structures of common autoinducers.
V. fischeri produces light within the light organ of squid as the population density reaches a critical threshold. This light helps the cephalopod avoid predation at night, which benefits the bacteria in turn (Graf and Ruby 1998). LuxI produces the autoinducer 3-oxo-C6-HSL (Table 1), which activates LuxR to induce luxCDABE transcription (Engebrecht et al. 1983; Engebrecht and Silverman 1984). This leads to luciferase synthesis and bioluminescence. A positive feedback loop maintains bioluminescence via increasing acyl-HSL levels (Moré et al. 1996). At low cell densities (LCDs), LuxI synthesizes C8-HSL (Figure 1), delaying autoinduction to conserve energy (Callahan and Dunlap 2000; Eberhard 1972; Kuo et al. 1996; Nealson et al. 1970). The QS system of many Gram-negative bacteria is highly specific, although homologous to the LuxIR type. A particular AHL molecule is only detected by the species that produced it (Watson et al. 2002).
Regulation of bioluminescence in V. fischeri. At LCD, luxI synthesis of C8-HSL would compete to inhibit the binding of 3-oxo-C6-HSL to LuxR. When the cell density reaches a threshold, it is able to produce enough 3-oxo-C6-HSL to combine with LuxR to form a complex, thereby initiating the transcription of luxCDABE and forming bioluminescence. Small ellipses represent signaling molecules and large ellipses represent proteins. HCD, high cell density; HSL, homoserine lactone; LCD, low cell density.
Pseudomonas aeruginosa primarily infects immunocompromised individuals, such as patients with AIDS, cystic fibrosis (CF), and urinary tract infections (Azam and Khan 2019). The pathogenicity of P. aeruginosa is linked to virulence factors regulated by a unique QS system, which involves two sets of AHL-mediated signaling circuits and a pathway mediated by 2-alkyl-4-quinolones (Latifi et al. 1996; Pesci et al. 1999) (Figure 2). LasI in P. aeruginosa produces 3-oxo-C12-HSL (Table 1), which binds to LasR, triggering a feedback loop that enhances the expression of LasI (Gambello and Iglewski 1991; Passador et al. 1993; Pearson et al. 1994; Schuster and Peter Greenberg 2006; Venturi 2006). This means that concentration of 3-oxo-C12-HSL is closely associated with P. aeruginosa's virulence. The second QS system consists of RhlI and RhlR, with RhlI synthesizing N-butanoyl-l-homoserine lactone (C4-HSL) (Table 1) (Pearson et al. 1995; Winson et al. 1995). The RhlR-HSL complex also forms a positive feedback loop, and these two signaling systems interact in sequence (M. B. Miller and Bassler 2001). The RhlIR system relies on the LasIR system for activation (Latifi et al. 1996; Pesci et al. 1997). Both systems regulate the production of specific virulence factors, including the elastin hydrolyzing protein LasB (Gambello and Iglewski 1991; Passador et al. 1993; Pearson et al. 1994; Pearson et al. 1995; Pearson et al. 1997; Winson et al. 1995). In addition, Pesci et al. identified a third signaling molecule, 2-heptyl-3-hydroxy-4-quinolone, referred to as the Pseudomonas quinolone signal (PQS) (Table 1). This signaling molecule is produced by the pqsABCDE operon and binds to PqsR to regulate the expression of traits (Cao et al. 2001; Pesci et al. 1999). The Las and Rhl systems exert positive and negative regulatory effects on the PQS system, respectively (Brouwer et al. 2014; Gallagher et al. 2002). Overall, P. aeruginosa's QS system is more complex than that of V. fischeri. The three systems are interconnected and collectively regulate the expression of virulence factors (Figure 2). Rhl and Pqs systems can regulate pyocin detected in CF patients (Gallagher et al. 2002; Mukherjee et al. 2017). All three systems participate in the generation of LasB (Brint and Ohman 1995; Diggle et al. 2003; Pesci et al. 1997). Bacteriocin and LasB play important roles in the pathogenesis of P. aeruginosa (S. Hall et al. 2016; Kuang et al. 2011), which also leads to the prevalence of P. aeruginosa in hospitals (Weiner et al. 2016). This complexity is a focus of ongoing research, particularly regarding the relationship between QS and drug resistance (Reynolds and Kollef 2021).
QS regulatory system in Pseudomonas aeruginosa. The yellow section represents the Rhl system. The blue part represents the Las system. The red part represents the Pqs system. The black arrow indicates upregulation and the red blocking arrow indicates downregulation. The small ellipses represent signaling molecules. The large ovals represent proteins. The Las system activates the Rhl and Pqs systems. However, the Rhl system has an inhibitory effect on the Pqs system. The three systems interact to express the production of various virulence factors. HSL, homoserine lactone; PQS, Pseudomonas quinolone signal; QS, quorum sensing.
Bacteria utilize QS circuits with high specificity to distinguish themselves from other species, enabling coordinated behaviors within bacterial communities. In addition to self-recognition, bacteria possess mechanisms to detect other species. V. harveyi possesses three QS systems for inter- and intraspecific communication that produce distinct signal molecules (Figure 3). Among them, N-(3-hydroxybutanoyl)-l-homoserine lactone (HAI-1) facilitates intraspecies communication (Bassler et al. 1993), while (S)-3-hydroxytridecan-4-one (CAI-1) operates within the Vibrio genus, and AI-2 is detectable by a broad range of bacteria, enabling cross-species signaling (Table 1) (Bassler et al. 1994). These systems involve Lux proteins in phosphorylation and dephosphorylation cascades that regulate LuxR and AphA (Ball et al. 2017; Freeman and Bassler 1999b; Freeman et al. 2000). AphA and LuxR are the major QS-regulated transcription factors (Rutherford et al. 2011). Among them, LuxR is only responsible for transcription rather than acting as a signaling receptor (Showalter et al. 1990). At an LCD, sRNAs inhibit LuxR (Lenz et al. 2004; Lilley and Bassler 2000), while at a high cell density (HCD), LuxR expression leads to bioluminescence (Freeman and Bassler 1999a; Lilley and Bassler 2000). This complex QS network underscores the versatility of bacterial communication, benefiting multispecies survival in shared environments (Liu et al. 2022).
Vibrio harveyi reacts to three autoinducers. HAI-1, AI-2, and CAI-1 are replaced by blue triangles, purple rectangles, and red circles, respectively. The large ovals represent proteins. The cylinders H and D denote conserved histidine and conserved aspartate, respectively. At LCD, LuxN, and LuxQ undergo autophosphorylation, sequentially transferring phosphate to LuxU and LuxO. Qrr-sRNA was activated to express AphA and inhibit LuxR. AphA at high levels is a major regulator at LCD. At HCD, LuxN, and LuxQ switch from kinases to phosphatases. Inactive LuxO is not phosphorylated. Disinhibition of LuxR by sRNA leaves LuxR at a high level. LuxR binds to the luxCDABE promoter, activating transcription. So, V. harveyi emits light. Double-headed arrows indicate the direction of transfer of phosphorylation. Phosphorylation shifted toward LuxO at LCD and the opposite at HCD. AI-2, autoinducer-2; CAI-1, (S)-3-hydroxytridecan-4-one; HAI-1, N-(3-hydroxybutanoyl)-l-homoserine lactone; HCD, high cell density; LCD, low cell density; QS, quorum sensing; sRNA, small RNA.
Beyond V. fischeri, P. aeruginosa, and V. harveyi, diverse LuxIR-based QS systems have been described (Table 2). For example, Agrobacterium tumefaciens induces crown gall disease through horizontally transferring T-DNA from virulence tumor-inducing (Ti) plasmids to plants, activating its QS system (Chevrot et al. 2006; Chilton et al. 1977; Haudecoeur and Faure 2010). A. tumefaciens' QS system regulates the spread of Ti plasmid with an N-(3-oxooctanoyl)-l-homoserine lactone signal (H. B. Zhang et al. 2002). Pseudomonas aureofaciens produces phenazine antibiotics, giving it a competitive edge against fungi and enabling it to serve as a biocontrol agent (Mazzola et al. 1992). In the QS circuit of P. aureofaciens, N-hexanoyl-l-homoserine lactone is synthesized by PhzI and binds to PhzR to regulate the expression of target genes (Wood et al. 1997; Wood and Pierson 1996). Pseudomonas fluorescens 2P24 utilizes the PcoIR system to regulate colonization around the rhizosphere and protect plant roots from disease (X. Yu et al. 2022). Sinorhizobium meliloti regulates the production of succinoglycan and EPS II through the AHL synthetase SinI and the receptor ExpR, allowing it to invade the nodules of Medicago legumes to form a symbiotic relationship through nitrogen fixation (Gao et al. 2005).
The variety of AHLs highlights the ubiquity of QS systems in these bacteria. Research has shown that QS in Gram-negative bacteria involves not only LuxI/R circuits but also two-component systems, as seen in Serratia (Van Houdt et al. 2007). As mentioned above, a wide variety of QS systems in Gram-negative bacteria are closely associated with virulence factors and pathogenicity. Thus, regulating QS to alter, reduce, or even inhibit the production of bacterial pathogenicity is a promising research direction (Landman et al. 2018).
3 Gram-Positive Bacteria
Gram-positive bacteria utilize self-inducing peptides (AIPs) for QS, unlike Gram-negative bacteria, which rely on homoserine lactones (HSLs). In Gram-negative bacteria, AHL signaling molecules usually activate responses by diffusing and binding to LuxR homolog receptors. By contrast, the QS system of Gram-positive bacteria includes a two-component system with a phosphorylation cascade and Rap, Rgg, NprR, PlcR and PrgX family that forms intracellular complexes. Signal transduction relies on the switch between the phosphorylation and dephosphorylation of sensor proteins to guide cascade reactions. (Neiditch et al. 2017). AIPs require exporters for release, ensuring specificity with sensor kinases and receptors (Table 3) (Waters and Bassler 2005).
Table 3.
List the QS systems of common Gram-positive pathogens and their autoinducers.
Abbreviations: AI-2, autoinducer-2; AIPs, self-inducing peptides; QS, quorum sensing.
Among Gram-positive bacteria, Staphylococcus aureus is most closely associated with human diseases. S. aureus causes disease by producing toxins and enzymes that damage tissues and lead to infections, especially in people with weakened immune systems (Lowy 1998; Turner et al. 2019). The Agr system, part of 16 two-component systems in S. aureus (Bleul et al. 2021), is central to the regulation of virulence (Matsumoto et al. 2021) (Figure 4). The Agr two-component system in S. aureus consists of transcription units driven by two promoters, P2 and P3. The system functions by producing a signal peptide, AIP, which is modified and exported outside the cell. The approximately 45-peptide remnant AgrD is processed, cyclized, and exported as AIP by the transmembrane endopeptidase AgrB. Once AIP reaches a threshold level, it binds to the histidine kinase AgrC, phosphorylating AgrA, which in turn regulates the P2 and P3 promoters to induce gene expression. This triggers a cascade that increases AIP levels, ensuring that an HCD is maintained (Reynolds and Wigneshweraraj 2011). RNAIII facilitates the production of the toxin α-hemolysin. During the activation of the agr operon, adhesin molecules are downregulated, while biofilm-degrading enzymes are upregulated. These shifts in gene expression enable S. aureus to transition from an adherent state to dispersion and proceed to its virulence phase (Dastgheyb et al. 2015; Novick et al. 1993). In addition to the Agr system, other two-component systems such as Sae, Sar, and Srr can influence S. aureus virulence through either affecting the Agr system or directly regulating the expression of virulence factors. As a result, the Agr system is subject to modulation by various environmental factors (Morrison 2012; Pragman et al. 2004; Voyich et al. 2009). Changes in the local environment during S. aureus infection can influence its pathogenicity (Mayville et al. 1999; Novick 2003). Therefore, when regulating virulence through QS, it is essential to account for the complexity of environmental variations.
Two-component sensor system for Staphylococcus aureus. P2 and P3, respectively, represent the promoters of operon agrBDCA and RNAIII transcription. Small circles with lactone ring structure indicate AIP produced by AgrB processing AgrD propeptide, and different colors represent AIP of different Staphylococcal strains origin. AIPs, self-inducing peptides.
Research has revealed the complexity of Gram-positive QS systems. The infection caused by S. pneumoniae, the leading source of community-acquired pneumonia, is closely linked to QS. Among the most extensively studied systems is the ComABCDE system, which resembles the Agr system of S. aureus (Table 3) (Karlsson et al. 2007). This system facilitates biofilm formation and bacteriocin production (Shanker and Federle 2017). In addition, the Com system is associated with horizontal gene transfer, a critical factor in S. pneumoniae pathogenesis and antibiotic resistance (Marks et al. 2012). Research by Domenech et al. explored disrupting the Com system to mitigate S. pneumoniae resistance (Domenech et al. 2020). Beyond the Com system, S. pneumoniae also possesses the BlpABCSRH and LuxS/AI-2 systems. The BlpABCSRH system regulates the production of class II bacteriocins and immunity proteins (Knutsen et al. 2004; Reichmann and Hakenbeck 2000), while the LuxS/AI-2 system influences biofilm formation on abiotic surfaces and is linked to upper respiratory tract infections (Vidal et al. 2011). Studies suggest that all three systems are involved in fratricide, a behavior that enables S. pneumoniae to suppress competing bacteria (Dawid et al. 2007; Steinmoen et al. 2002). Furthermore, increasing evidence indicates that the LuxS/AI-2 system is not exclusive to S. pneumoniae but is also widely present in other Gram-positive bacteria, such as S. aureus (D. Yu et al. 2012), Bacillus anthracis (Bozue et al. 2012), Borrelia burgdorferi (Stevenson and Babb 2002), Clostridium difficile (Carter et al. 2005), and Enterococcus faecalis (L. Ali, Goraya, et al. 2017) (Table 3). The AI-2 molecule significantly influences biofilm formation (Vinodhini and Kavitha 2024), cell adhesion, virulence, and antibiotic resistance (Y. Wang et al. 2019) in Gram-positive bacteria. Because AI-2 serves as an interspecies signaling molecule, blocking these processes to influence the virulence of these pathogens may present a novel research direction. It is noteworthy that the QS systems of both Gram-negative and Gram-positive bacteria often regulate downstream phenotypes such as virulence and biofilms through multiple mechanisms, rather than a single circuit acting in isolation (Miyamoto et al. 2000).
4 Quorum Sensing Quenching and Human Disease Treatment
As the QS system serves as a crucial communication pathway for pathogenic bacteria that threaten human health, blocking QS to protect human health is therefore a vital research direction. The applications of QS include the use of biosensors for bacterial detection, the disruption of biofilm formation to prevent antibiotic resistance, and the development of potential antitumor strategies.
4.1 Quorum Quenching (QQ) and Quorum Sensing Inhibitors
Since the 1940s, antibiotic overuse has spurred microbial resistance to antibiotics. The intense selection pressure brought about by excessive antibiotic usage has led to the continuous emergence of drug-resistant strains (Bell et al. 2014). Selection pressure caused by antibiotics is unavoidable, necessitating the frequent development of new antibiotics (Kalia et al. 2007). QQ offers a resistance-free solution through targeting QS systems to reduce bacterial pathogenicity (Paluch et al. 2020).
Generally, QS inhibitors (QSIs) lead to QQ through three main mechanisms. The first is targeting signaling molecules (Anandan and Vittal 2019; Chung et al. 2011). This mechanism includes the inhibition of the production of signaling molecules (Hirakawa and Tomita 2013) and the degradation of signaling molecules by QQ enzymes. Between the two, the most well-known mechanism is the degradation of signaling molecules. Typical QQ enzymes include lactonases, acyltransferases, oxidases and reductases (Rehman and Leiknes 2018). QQ enzymes degrade signaling molecules, preventing their accumulation to threshold levels and thereby effectively blocking the onset of QS. The second mechanism targets the receptors of signaling molecules. This involves using antagonists to competitively block the binding of signaling molecules to their receptors (Bodede et al. 2018; Proctor et al. 2020). The final mechanism involves disrupting the downstream signaling cascade. This is primarily achieved through kinase inhibition (Brackman and Coenye 2015) and interference with the transcriptional regulators of operon genes (Figure 5) (Sully et al. 2014).
The mechanism of the three QSI. On the left are Gram-negative bacteria. Triangles represent AHL signaling molecules. Rectangles represent receptor antagonists. Ellipses represent proteins. On the right are Gram-positive bacteria. Red circles represent AIP, and blue circles represent AIP antagonists. Rectangles represent kinase inhibitors. The pink ellipse represents the propeptide, and the purple ellipse represents the protein. (i) Targeting signaling molecules. (1) Inhibits the synthesis of signaling molecules. (2) Degradation of signaling molecules to prevent aggregation. (ii) Targeting signaling molecule receptors. (3) Competitively binding receptors. (iii) Interference with downstream signaling cascades. (4) Prevents kinase sensor activation. (5) Blocking the binding of AgrA to DNA. AHL, acyl-homoserine lactone; AIPs, self-inducing peptides; QSI, quorum sensing inhibitor.
The applications of QQ are diverse and promising. V. harveyi, a major marine pathogen, threatens shrimp and causes economic losses in aquaculture (Sheikh et al. 2022). The rise of antibiotic resistance underscores the potential of targeting the QS system to combat this issue. Research has shown that AI-2 signaling molecules in V. harveyi's QS system play a crucial role in regulating virulence in brine shrimp (Defoirdt et al. 2005). Inactivating LuxS or LuxP disrupts shrimp mortality (Defoirdt et al. 2008). Furanones such as (5Z)-4-bromo-5-(bromomethylene)-3-butyl-2(5H)-furanone can block LuxR (Defoirdt et al. 2007). This way of interfering with the QS circuit does not increase the selective pressure to keep more resistant strains alive as with antibiotics, and thus can reduce pathogenicity without developing resistance (J. Chen et al. 2020). In addition, QQ can inhibit the luminescence of V. harveyi and the expression of toxin T1, thereby reducing the infectivity of V. harveyi to shrimp and its impact on aquaculture (Manefield et al. 2000). Reducing the AHL signal concentration using the AiiA enzyme can inhibit the virulence of Erwinia carotovora and thus improve the soft rot resistance of common vegetables, such as cabbage and potato (Dong et al. 2000). The discovery of farnesol in Candida albicans has extended the QS range to fungi (Hornby et al. 2001). Increasing the concentration of farnesol can inhibit the formation of Candida biofilm and decrease the resistance (Zibafar et al. 2009). Considering the drug resistance issues associated with antibiotic misuse, the innovative mechanism of QQ holds broader potential, making it an exciting and promising research avenue (Table 4). However, clinical QQ application demands rigorous validation due to potential toxicity and resistance risks (Bjarnsholt and Givskov 2007; Maeda et al. 2012), necessitating thorough safety assessments and strategies to address ecological and practical challenges (Maddela et al. 2019). Furthermore, research suggests that employing QQ to treat bacterial infections may lead to issues similar to antibiotic resistance (Defoirdt 2018), which contrasts with the ideal scenario outlined above. Given these limitations, future research on QQ and QSI must aim for a deeper understanding and resolution of these issues.
Table 4.
Advances in research to suppress pathogenicity by blocking quorum sensing.
Bacteria
Inhibitor
Function
Reference
Inhibit the synthesis of AHLs
Pseudomonas aeruginosa
Enoyl-acyl carrier protein reductase inhibitors
Inhibition of enoyl-acyl carrier protein reductase in the bacterial type II fatty acid synthesis pathway
Reduce the 3-oxo-C12-HSL molecules; interferes with the synthesis of the autoinducers itself by reducing the free 3-oxo- acyl carrier protein in the cell
Abbreviations: AHL, acyl-homoserine lactone; HSL, homoserine lactone; QQ, quorum quenching; QS, quorum sensing.
4.2 Biosensor and Human Disease Prevention
QS-based biosensors can rapidly and accurately assess bacterial populations in diverse settings, including clinical, food production, and environmental monitoring contexts. By detecting QS molecules, these biosensors enable the early detection of bacterial contamination, facilitating timely interventions to prevent infections and spoilage (C. Miller and Gilmore 2020; Paluch et al. 2020).
Specifically, biosensors need to be highly sensitive to HSL signals in the QS circuits of pathogenic bacteria, detecting specific pathogens via homologous binding proteins, such as LuxR. Water quality is crucial for human health, making biosensors valuable for the efficient detection of waterborne pathogens. Researchers have developed sensor modules, such as QscR, for detecting bacterial contamination in water (Wu et al. 2021). In the field of food safety, biosensors can be employed to monitor food decay based on luminescence responses to volatile organic compounds, although further development is needed to enhance the QS signal response (Veltman et al. 2022). The V. harveyi-based whole-cell biosensor can detect the AI-2 signaling molecule expressed by Campylobacter jejuni, which reduces the detection limit by 100 times and can be employed for quantitative detection and the improvement of food production safety (Ramić et al. 2022). Whether the application of QS circuits or homologous binding proteins can be influenced to improve the universality in future studies is a key question. The application of QS sensors can be widely utilized for bacterial detection in food and clinical settings, with profound implications for the prevention and treatment of human diseases (Fan et al. 2022).
4.3 Biofilm and Resistance
Because traditional antimicrobial drugs directly target bacteria, bacterial resistance is easily induced in environments with high selection pressure (Jantaruk et al. 2021). QS-based strategies disrupt communication, reducing virulence without killing bacteria, thereby potentially enhancing antibiotic efficacy and offering a promising route for next-generation treatments (Paluch et al. 2020; X. Yu et al. 2021). In addition, the ability to modulate QS signaling could potentially enhance the effectiveness of existing antibiotics, making them more potent against antibiotic-resistant bacterial strains (Rogers et al. 2012). Thus, leveraging QS in biosensors and therapeutic interventions presents a multifaceted approach to combat bacterial resistance and improve public health outcomes.
Biofilms are complex and highly structured communities of microorganisms that attach to surfaces (Costerton et al. 1995). Biofilms can grow on a variety of surfaces, both living and nonliving, such as tissues and organs, medical devices, teeth, and rocks (Li and Zhao 2020). Biofilms provide numerous advantages to microorganisms, including protection from environmental stressors (e.g., antibiotics and immune responses), enhanced nutrient availability, and a platform for cooperative behaviors (Miquel et al. 2016). Biofilm formation is a gradual process that involves successive stages, namely, bacterial adhesion to the host surface, biofilm accumulation, maturation, and eventual detachment (Stoodley et al. 2002).
When bacteria encounter a suitable surface for survival, they can attach to a surface to form a biofilm. Bacterial biofilms are extremely important (Kolter and Greenberg 2006), being closely related to pathogenicity (Woelber et al. 2022), resistance (C. W. Hall and Mah 2017), and biotechnology (Edel et al. 2019). Studies have shown that more than 80% of human pathogen infections are associated with biofilm formation (Bryers 2008). Bacteria transmit attachment signals through QS. Different bacteria exhibit variations in biofilm formation. For example, P. aeruginosa forms biofilms at an HCD, whereas Vibrio cholerae and S. aureus form biofilms at an LCD (Bronesky et al. 2016; de Kievit and Iglewski 2000; Waters et al. 2008). QS plays an important role in the biofilm formation of both Gram-positive and Gram-negative bacteria. The expression of genes responsible for biofilm formation can be controlled by regulating bacterial QS (Khan et al. 2019). Once these QS circuits are affected, bacterial colonization, adhesion, and the production of virulence factors are greatly reduced (Y. Zhang et al. 2018). Biofilms provide pathogens with a physical barrier that can resist the host's immune system and external adverse factors. Therefore, biofilm formation is crucial for the development of drug resistance. Using small molecules to block the QS circuit to affect biofilm formation is considered a potential strategy for reducing bacterial pathogenicity.
The QS system in P. aeruginosa, particularly the Las and Rhl system, plays a crucial role in regulating biofilm formation through influencing virulence factors, such as rhamnolipids (Smit 2003). Research indicates that rhamnolipids controlled by the Rhl system can impact biofilm aggregation (Tielker et al. 2005; Winzer et al. 2000), with mutant strains exhibiting different biofilm structures compared with wild-type strains (Passos da Silva et al. 2017). Various factors such as AHL analogs (Hentzer et al. 2002), lectins such as LecA and LecB (Tielker et al. 2005; Winzer et al. 2000), and iron siderophores (Banin et al. 2005) also play roles in biofilm formation. The PQS system increases extracellular DNA content, interacting with extracellular polymeric substances to form biofilms (Allesen-Holm et al. 2006; Diggle et al. 2003; Yang et al. 2007). In addition, the luxS gene has been linked to biofilm formation in various bacteria, including S. aureus (Xue et al. 2013), Helicobacter pylori (Cole et al. 2004), and Streptococcus mutans (Merritt et al. 2003), in which it has inhibitory effects on biofilm formation.
In summary, QS regulates bacterial detachment from mature biofilms, enabling their dispersion to new sites for colonization and biofilm formation. This QS-biofilm interplay is a compelling area of research, highlighting bacterial communicate, and environmental adaptation. Reducing biofilm formation is crucial for developing bacteria-resistant medical devices (Paluch et al. 2018) and antisepsis and anti-biofilm metals (Piecuch et al. 2016), as well as for disrupting bacterial virulence, with broad implications for combating biofilm-related infections and controlling bacterial growth in industrial systems.
4.4 Cancer
Cancer is a critical noncommunicable disease (Bray et al. 2018) that necessitates the development of innovative therapies due to treatment resistance. The use of bacterial-mediated cancer therapy and QS to develop tumor-targeted drug-killing tools can minimize side effects (Patel and Spassieva 2018). Studies have shown that the specific microenvironment in necrotic tissues such as tumors can attract the preferential aggregation of Salmonella (Forbes et al. 2003). By utilizing this characteristic, the selective expression of therapeutic proteins activated by QS after the accumulation of Salmonella in tumors can minimize damage to healthy tissues (M. Zhang et al. 2014). This approach overcomes the limitations of conventional therapies and does not rely on tumor surface markers. Salmonella at threshold concentrations in tumors triggers QS gene expression, selectively releasing therapeutic proteins within the cancer environment (Pawelek et al. 2003). Aganja et al. enhanced Salmonella's anticancer potency by detoxifying, attenuating virulence, and utilizing QS for tumor-targeted toxin expression, thus avoiding damage to healthy tissues (Aganja et al. 2024). The specific expression range and limited triggering factors allow Salmonella to minimize side effects (Ganai et al. 2009; Loessner et al. 2007).
In addition to Salmonella, many additional bacteria, such as Cyanobacteria (Liang et al. 2021) and Enterococcus faecium (Wynendaele et al. 2022), have also been found to hold promise for cancer therapy. In conclusion, the specific expression of drug proteins by QS can avoid side effects while exerting cancer control. However, several challenges remain, including preventing the spread of bacterial infections after antitumor activity (Forbes et al. 2018), achieving the highest possible bacterial targeting to improve specificity, and managing various virulence factors induced by bacterial QS or immune responses triggered by the bacteria itself (Kalia et al. 2022). As a result, most studies utilize attenuated or detoxified strains that have been screened to ensure the health of the organism (Kalia et al. 2022; Kumar et al. 2022).
Today, humanity faces the critical challenge of rising antibiotic ineffectiveness (Kwon and Powderly 2021). However, research on QQ enzymes and QSI offers new hope for overcoming this predicament. Blocking biofilm formation and expression of virulence factors by interfering with the QS system may avoid resistance resulting from the survival of resistant strains under selective pressure. This nonselective pressure strategy holds promise as a potential antibiotic alternative (Kalia et al. 2019) and offers a new avenue for addressing the challenges of treating cancer (Liang et al. 2021). Whether in clinical applications (Borges and Simões 2019; Carradori et al. 2020) or industrial settings (Defoirdt et al. 2011; Skindersoe et al. 2008), QS regulatory technologies have demonstrated significant potential. However, compared with controlled laboratory conditions, the bacterial communities and QS networks in real-world environments are far more complex. Further research is essential to enhance the specificity and targeting accuracy of intricate QS systems, paving the way for more effective and safe applications.
5 Novel QS Regulation Mode
The greater the harm that bacteria pose to human health, the more significant QS becomes in disease prevention and control. Currently, many novel and unexpected QS regulatory mechanisms have been discovered. For example, new materials such as nanomaterials and hydrogel materials can be used in combination with QS. In addition, to protect the environment and human health through QS degradation of microplastics (MPs) by bacteria. Furthermore, the use of phages to protect humans is also a promising direction. These new solutions will greatly improve the application scope and application value of QS.
5.1 Nanomaterial Regulation
We are currently in the postantibiotic era. QS techniques hold promise in mitigating bacterial virulence without triggering resistance, making research on QSI and QQ enzymes highly active. However, recent concerns have been raised regarding the low bioavailability and high toxicity of QSI (Gómez-Gómez et al. 2019). Nanomaterials, with their superior properties, not only have the potential to directly modulate QS but can also significantly enhance the efficiency and stability of QSI. Therefore, nanocomposites make QQ more feasible.
Nanomaterials are widely utilized in various applications. Depending on their distinct properties, nanomaterials can serve as optical sensors (Vasudevan et al. 2021), electrochemical sensors (Capatina et al. 2022), and bacterial biosensors (Guan et al. 2018), enabling the detection and diagnosis of diseases when bacterial populations reach a certain threshold. Nanomaterials can prevent bacterial colonization and spread (Roy et al. 2022) and inhibit the secretion of virulence factors (Lu et al. 2020). In addition, nanomaterials can disrupt and inhibit biofilm formation. This results in a significant reduction in bacterial virulence and defense mechanisms, making prevention and treatment more straightforward and effective (Kamli et al. 2021).
Nanomaterials primarily interfere with QS through inhibiting the supply and transduction of signaling molecules (Figure 6). In terms of the signal supply, nanomaterials can block the supply of precursor molecules (A1) (Shah et al. 2019), alter the conformation of enzymes such as LuxI synthase (A2) (S. G. Ali et al. 2017), or block the active sites of these enzymes (A3) (Shah et al. 2019). Nanomaterials also prevent the secretion of signaling molecules into the extracellular space via inducing aggregation effects (A4) (Omwenga et al. 2018) and suppressing efflux pumps (A5) (Ahmed et al. 2021). In addition, nanomaterials can adsorb signaling molecules due to their adsorptive properties (A6) (Piletska et al. 2010) or catalyze the degradation of signaling molecules (A7) (Jegel et al. 2021). This prevents the extracellular accumulation of QS signals, hindering their ability to reach the threshold necessary to trigger the expression of virulence factors. In terms of signal transduction, nanomaterials can competitively bind to cytoplasmic and membrane receptors (B1, B2) (Balakrishnan et al. 2020; Omwenga et al. 2018), disrupting the functionality of the cell membrane. In addition, nanomaterials can even directly damage DNA to inhibit gene expression (B3) (L. Wang et al. 2017).
Mechanisms of nanomaterial regulation in QS. The green box (A1–A7) represents signal supply inhibition, while the purple box (B1–B3) represents signal transduction inhibition. The left side illustrates Gram-negative bacteria, and the right side illustrates Gram-positive bacteria. See the text for detailed mechanisms. AHL, acyl-homoserine lactone; AIPs, self-inducing peptides; QS, quorum sensing.
However, due to constraints in nanomaterial processing and delivery methods, the current application of nanomaterials to regulate QS is primarily limited to treating diseases of the digestive and respiratory systems (Mogosanu et al. 2015). Expanding nanomaterial modification techniques could greatly enhance their therapeutic value. For example, injectable nanomaterials could be utilized to treat bloodborne or systemic bacterial infections (Peng et al. 2024), or topical applications could be absorbed through the skin to treat bone or joint disorders (Cheng et al. 2022). This expansion would require focused research on nanomaterial dosage and safety. In addition, nanomaterials currently face issues with stability, which presents challenges in storage and transportation. Most importantly, the microbial environment in the human body is far more complex than that in a laboratory setting, necessitating extensive experimentation to validate the effectiveness of nanomaterials.
5.2 Hydrogel Regulation
Hydrogels are an intriguing material that has garnered increasing research interest in recent years. Currently, the design of hydrogels allows them to respond to a variety of environmental changes. For example, hydrogels can release specific substances in response to stimuli, such as temperature, pH, or light, thereby regulating or inhibiting bacterial QS. This represents a novel approach for treating many infectious diseases.
Research on utilizing hydrogels to regulate QS across various dimensions has highlighted its advantages, such as the inhibition of biofilm formation (Hu et al. 2022; Khaleghi et al. 2023), the suppression of virulence factor production (Prateeksha et al. 2019), anti-inflammatory properties (Tu et al. 2022), anti-infection efficacy (Qu et al. 2024), detection capabilities (Seto 2019), and applications in food packaging and seafood preservation (D. Wang et al. 2023). For example, high concentrations of reactive oxygen species (ROS) produced by bacterial infections can exacerbate inflammation, thereby hindering the treatment of posttraumatic osteomyelitis (W. Zhang et al. 2024). Zhang et al. synthesized a hydrogel that releases hyperbranched polylysines (HBPLs) in response to changes in ROS and pH levels in the environment. Under acidic conditions and in the presence of high H2O2 concentrations, the hydrogel releases HBPL more rapidly. HBPL effectively scavenges ROS, disrupts bacterial biofilm formation, and interferes with the expression of QS-related genes (W. Zhang et al. 2024). In addition, Seto developed a hydrogel capable of detecting P. aeruginosa infections in a cell-free environment. This cell-free compatible hydrogel demonstrates very high sensitivity and practicality (Seto 2019).
Due to their unique viscoelasticity and rheological properties, hydrogels are widely employed in the field of delivery systems. This suggests that incorporating stimuli-responsive agents and therapeutic substances such as QSI into hydrogels could enable the disruption of QS, thereby achieving therapeutic outcomes. However, research on the use of hydrogels for QS regulation remains limited, and whether hydrogels can realistically serve as an alternative to antibiotics, offering a novel therapeutic approach, requires extensive experimental validation in further research.
5.3 QS and MP Degradation
MPs are defined as plastic particles or fragments with a diameter of less than 5 mm (Thompson et al. 2004). Today, MPs are distributed globally (Ivar do Sul et al. 2009) and pose serious risks to organisms within the environment (Fueser et al. 2020). As apex predators in the food chain, humans can accumulate significant amounts of MPs in their bodies through biomagnification, leading to potential harm (Barboza et al. 2018; Vethaak and Legler 2021). Leveraging bacterial QS holds promise for addressing this issue.
Some bacteria utilize MPs as a transport vector, attaching to the surface of MPs to form biofilms and create a plastisphere, which enables the long-distance transport of microorganisms (Zettler et al. 2013). The plastisphere is a miniature ecosystem that can harbor a variety of microorganisms (Amaral-Zettler et al. 2020). As a result, MPs facilitate the diffusion of microorganisms and influence aquatic ecosystems (Ahmad et al. 2020). Rhodobacteraceae are abundant in the plastisphere, likely due to the crucial regulatory role of conserved QS signals in biofilm formation (Debroas et al. 2017; Zan et al. 2014). As mentioned earlier, targeting QS to degrade MPs may represent a key research direction. Bacillus cereus has been shown to play an important role in the degradation of polyethylene, polyethylene terephthalate, and polyethylene terephthalate (X. Xu et al. 2023). As demonstrated by Su et al. through experimentation, QS may be a significant factor influencing the degradation of marine particulate matter (Su et al. 2018). However, research on the QS-mediated degradation of MPs remains limited and lacks comprehensive understanding. Because QS can enhance the bacterial recognition and degradation of MPs, introducing QS signaling molecules or analogs to activate bacterial degradation pathways represents another promising research direction. Furthermore, employing genetic engineering and other means of modifying the QS system to enable the expression of additional MP-degrading enzymes is a promising approach. In conclusion, the use of bacterial QS systems will ameliorate the effects of MPs on the marine environment. More importantly, reducing the threat of MP has a great effect on human health.
5.4 Virus Monitoring
Given the increasingly serious issue of antimicrobial resistance and a growing body of research on interkingdom QS, bacteriophages have received increasing attention (Luong et al. 2020). Some phages lay dormant until activated by signals from the bacterial communication system. Virulent phages (Suramo et al. 1981) replicate and destroy bacteria, while temperate phages (Hay and Lithgow 2019) can either kill the host (Ofir and Sorek 2018) or coexist through lysogeny (Lwoff 1953). Polylysogeny allows multiple prophages in a bacterium, but DNA damage can disrupt this balance (Campbell et al. 2020). Interestingly, DNA damage does not always induce phage replication. The host response to infection and QS signals can trigger the transition from lysogenic to lytic cycles in temperate phages (Silpe et al. 2023). Phages can also “monitor” bacterial signals to determine the best time to replicate. Elucidating this process can potentially enable the use of QS regulation to induce bacteriophages to eliminate harmful bacteria, presenting a novel approach to combat infections. This approach can avoid the selective pressure caused by drugs, which gives rise to bacterial resistance. Frequent antibiotic changes are costly. Therefore, the use of bacteriophages is expected to replace antibiotics to change this situation (Jin et al. 2023). It is noteworthy that phages can monitor specific autoinducers expressed by bacteria while also being triggered for dissemination (Silpe and Bassler 2019). This means that improper control could lead to phage infection following bacterial lysis. Therefore, the application of phages requires extensive research to identify a balanced state among bacteria, viruses, and the human body. This is essential to ensure that harmful bacteria can be monitored and eliminated without compromising health.
6 Conclusion and Perspective
QS is pivotal in bacterial communication, with progress in gene expression and regulatory mechanisms. It facilitates microbial interactions and adaptation across ecosystems, prompting further research into its cross-kingdom impacts. QS emerges as a strategic solution to pressing challenges, including microbial pathogenicity, antibiotic resistance, and cancer. It has the potential to mitigate pathogen virulence and regulate pathogenic bacteria through viral agents, reducing the risks of antibiotic overuse. The integration of QS with nanomaterials and hydrogels enhances its potential as a microbial control tool. However, the intricacies of QS networks and the variability in nonideal natural conditions present substantial challenges that necessitate further research and validation. In essence, QS has transcended its role as a communication mechanism to become an indispensable conduit for human modulation of microbial behavior.
This study was supported by the Natural Science Foundation of Shandong Province (ZR2022QC140) and State Key Laboratory of Microbial Technology Open Projects Fund (M2022-03).
Aganja, R. P., C. Sivasankar, and J. H. Lee. 2024. “AI-2 Quorum Sensing Controlled Delivery of Cytolysin-A by Tryptophan Auxotrophic Low-Endotoxic Salmonella and Its Anticancer Effects in CT26 Mice With Colon Cancer.” Journal of Advanced Research61: 83–100. https://doi.org/10.1016/j.jare.2023.09.003.
Ahmad, M., J.-L. Li, P.-D. Wang, W. N. Hozzein, and W.-J. Li. 2020. “Environmental Perspectives of Microplastic Pollution in the Aquatic Environment: A Review.” Marine Life Science & Technology2, no. 4: 414–430. https://doi.org/10.1007/s42995-020-00056-w.
Ahmed, F. Y., U. F. Aly, R. M. Abd El-Baky, and N. G. F. M. Waly. 2021. “Effect of Titanium Dioxide Nanoparticles on the Expression of Efflux Pump and Quorum-Sensing Genes in MDR Pseudomonas aeruginosa Isolates.” Antibiotics (USSR)10, no. 6: 625. https://doi.org/10.3390/antibiotics10060625.
Ali, S. G., M. A. Ansari, Q. M. Sajid Jamal, et al. 2017. “Antiquorum Sensing Activity of Silver Nanoparticles in P. aeruginosa: An In Silico Study.” In Silico Pharmacology5: 12. https://doi.org/10.1007/s40203-017-0031-3.
Ali, L., M. Goraya, Y. Arafat, M. Ajmal, J. L. Chen, and D. Yu. 2017. “Molecular Mechanism of Quorum-Sensing in Enterococcus faecalis: Its Role in Virulence and Therapeutic Approaches.” International Journal of Molecular Sciences18, no. 5: 960. https://doi.org/10.3390/ijms18050960.
Allesen-Holm, M., K. B. Barken, L. Yang, et al. 2006. “A Characterization of DNA Release in Pseudomonas aeruginosa Cultures and Biofilms.” Molecular Microbiology59, no. 4: 1114–1128. https://doi.org/10.1111/j.1365-2958.2005.05008.x.
Amaral-Zettler, L. A., E. R. Zettler, and T. J. Mincer. 2020. “Ecology of the Plastisphere.” Nature Reviews Microbiology18, no. 3: 139–151. https://doi.org/10.1038/s41579-019-0308-0.
Anandan, K., and R. R. Vittal. 2019. “Quorum Quenching Activity of AiiA Lactonase (KMMI17) From Endophytic Bacillus thuringiensis KMCL07 on AHL-Mediated Pathogenic Phenotype in Pseudomonas aeruginosa.” Microbial Pathogenesis132: 230–242. https://doi.org/10.1016/j.micpath.2019.05.015.
Azam, M. W., and A. U. Khan. 2019. “Updates on the Pathogenicity Status of Pseudomonas aeruginosa.” Drug Discovery Today24, no. 1: 350–359. https://doi.org/10.1016/j.drudis.2018.07.003.
Balaban, N., and R. P. Novick. 1995. “Autocrine Regulation of Toxin Synthesis by Staphylococcus aureus.” Proceedings of the National Academy of Sciences92, no. 5: 1619–1623. https://doi.org/10.1073/pnas.92.5.1619.
Balakrishnan, S., K. S. Ibrahim, S. Duraisamy, et al. 2020. “Antiquorum Sensing and Antibiofilm Potential of Biosynthesized Silver Nanoparticles of Myristica fragrans Seed Extract Against MDR Salmonella enterica serovar Typhi Isolates From Asymptomatic Typhoid Carriers and Typhoid Patients.” Environmental Science and Pollution Research27, no. 3: 2844–2856. https://doi.org/10.1007/s11356-019-07169-5.
Ball, A. S., R. R. Chaparian, and J. C. van Kessel. 2017. “Quorum Sensing Gene Regulation by LuxR/HapR Master Regulators in Vibrios.” Journal of Bacteriology199, no. 19: e00105-17. https://doi.org/10.1128/JB.00105-17.
Banin, E., M. L. Vasil, and E. P. Greenberg. 2005. “Iron and Pseudomonas aeruginosa Biofilm Formation.” Proceedings of the National Academy of Sciences102, no. 31: 11076–11081. https://doi.org/10.1073/pnas.0504266102.
Barboza, L. G. A., A. Dick Vethaak, B. R. B. O. Lavorante, A. K. Lundebye, and L. Guilhermino. 2018. “Marine Microplastic Debris: An Emerging Issue for Food Security, Food Safety and Human Health.” Marine Pollution Bulletin133: 336–348. https://doi.org/10.1016/j.marpolbul.2018.05.047.
Bareia, T., S. Pollak, and A. Eldar. 2018. “Self-Sensing in Bacillus subtilis Quorum-Sensing Systems.” Nature Microbiology3, no. 1: 83–89. https://doi.org/10.1038/s41564-017-0044-z.
Bassler, B. L., M. Wright, and M. R. Silverman. 1994. “Multiple Signalling Systems Controlling Expression of Luminescence in Vibrio harveyi: Sequence and Function of Genes Encoding a Second Sensory Pathway.” Molecular Microbiology13, no. 2: 273–286. https://doi.org/10.1111/j.1365-2958.1994.tb00422.x.
Bassler, B. L., M. Wright, R. E. Showalter, and M. R. Silverman. 1993. “Intercellular Signalling in Vibrio harveyi: Sequence and Function of Genes Regulating Expression of Luminescence.” Molecular Microbiology9, no. 4: 773–786. https://doi.org/10.1111/j.1365-2958.1993.tb01737.x.
Bell, B. G., F. Schellevis, E. Stobberingh, H. Goossens, and M. Pringle. 2014. “A Systematic Review and Meta-Analysis of the Effects of Antibiotic Consumption on Antibiotic Resistance.” BMC Infectious Diseases14: 13. https://doi.org/10.1186/1471-2334-14-13.
Bjarnsholt, T., and M. Givskov. 2007. “Quorum-Sensing Blockade as a Strategy for Enhancing Host Defences Against Bacterial Pathogens.” Philosophical Transactions of the Royal Society, B: Biological Sciences362, no. 1483: 1213–1222. https://doi.org/10.1098/rstb.2007.2046.
Bleul, L., P. Francois, and C. Wolz. 2021. “Two-Component Systems of S. aureus: Signaling and Sensing Mechanisms.” Genes13, no. 1: 34. https://doi.org/10.3390/genes13010034.
Bodede, O., S. Shaik, H. Chenia, P. Singh, and R. Moodley. 2018. “Quorum Sensing Inhibitory Potential and In Silico Molecular Docking of Flavonoids and Novel Terpenoids From Senegalia nigrescens.” Journal of Ethnopharmacology216: 134–146. https://doi.org/10.1016/j.jep.2018.01.031.
Bozue, J., B. S. Powell, C. K. Cote, et al. 2012. “Disrupting the luxS Quorum Sensing Gene Does Not Significantly Affect Bacillus anthracis Virulence in Mice or Guinea Pigs.” Virulence3, no. 6: 504–509. https://doi.org/10.4161/viru.21934.
Bray, F., J. Ferlay, I. Soerjomataram, R. L. Siegel, L. A. Torre, and A. Jemal. 2018. “Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries.” CA: A Cancer Journal for Clinicians68, no. 6: 394–424. https://doi.org/10.3322/caac.21492.
Brint, J. M., and D. E. Ohman. 1995. “Synthesis of Multiple Exoproducts in Pseudomonas aeruginosa Is under the Control of RhlR-RhlI, Another Set of Regulators in Strain PAO1 With Homology to the Autoinducer-Responsive LuxR–LuxI Family.” Journal of Bacteriology177, no. 24: 7155–7163. https://doi.org/10.1128/jb.177.24.7155-7163.1995.
Bronesky, D., Z. Wu, S. Marzi, et al. 2016. “Staphylococcus aureus RNAIII and Its Regulon Link Quorum Sensing, Stress Responses, Metabolic Adaptation, and Regulation of Virulence Gene Expression.” Annual Review of Microbiology70: 299–316. https://doi.org/10.1146/annurev-micro-102215-095708.
Brouwer, S., C. Pustelny, C. Ritter, B. Klinkert, F. Narberhaus, and S. Häussler. 2014. “The PqsR and RhlR Transcriptional Regulators Determine the Level of Pseudomonas Quinolone Signal Synthesis in Pseudomonas aeruginosa by Producing Two Different pqsABCDE mRNA Isoforms.” Journal of Bacteriology196, no. 23: 4163–4171. https://doi.org/10.1128/JB.02000-14.
Butrico, C. E., and J. E. Cassat. 2020. “Quorum Sensing and Toxin Production in Staphylococcus aureus Osteomyelitis: Pathogenesis and Paradox.” Toxins12, no. 8: 516. https://doi.org/10.3390/toxins12080516.
Calatrava-Morales, N., M. McIntosh, and M. J. Soto. 2018. “Regulation Mediated by N-Acyl Homoserine Lactone Quorum Sensing Signals in the Rhizobium-Legume Symbiosis.” Genes9, no. 5: 263. https://doi.org/10.3390/genes9050263.
Callahan, S. M., and P. V. Dunlap. 2000. “LuxR- and Acyl-Homoserine-Lactone-Controlled Non-Lux Genes Define a Quorum-Sensing Regulon in Vibrio fischeri.” Journal of Bacteriology182, no. 10: 2811–2822. https://doi.org/10.1128/JB.182.10.2811-2822.2000.
Campbell, D. E., L. K. Ly, J. M. Ridlon, A. Hsiao, R. J. Whitaker, and P. H. Degnan. 2020. “Infection With Bacteroides Phage BV01 Alters the Host Transcriptome and Bile Acid Metabolism in a Common Human Gut Microbe.” Cell Reports32, no. 11: 108142. https://doi.org/10.1016/j.celrep.2020.108142.
Cao, H., G. Krishnan, B. Goumnerov, J. Tsongalis, R. Tompkins, and L. G. Rahme. 2001. “A Quorum Sensing-Associated Virulence Gene of Pseudomonas aeruginosa Encodes a LysR-Like Transcription Regulator With a Unique Self-Regulatory Mechanism.” Proceedings of the National Academy of Sciences98, no. 25: 14613–14618. https://doi.org/10.1073/pnas.251465298.
Capatina, D., T. Lupoi, B. Feier, D. Olah, C. Cristea, and R. Oprean. 2022. “Highly Sensitive Detection of PQS Quorum Sensing in Pseudomonas aeruginosa Using Screen-Printed Electrodes Modified With Nanomaterials.” Biosensors12, no. 8: 638. https://doi.org/10.3390/bios12080638.
Carradori, S., N. Di Giacomo, M. Lobefalo, G. Luisi, C. Campestre, and F. Sisto. 2020. “Biofilm and Quorum Sensing Inhibitors: The Road so Far.” Expert Opinion on Therapeutic Patents30, no. 12: 917–930. https://doi.org/10.1080/13543776.2020.1830059.
Carter, G. P., D. Purdy, P. Williams, and N. P. Minton. 2005. “Quorum Sensing in Clostridium difficile: Analysis of a luxS-Type Signalling System.” Journal of Medical Microbiology54, no. Pt 2: 119–127. https://doi.org/10.1099/jmm.0.45817-0.
Chen, J., Y. Lu, X. Ye, M. Emam, H. Zhang, and H. Wang. 2020. “Current Advances in Vibrio harveyi Quorum Sensing as Drug Discovery Targets.” European Journal of Medicinal Chemistry207: 112741. https://doi.org/10.1016/j.ejmech.2020.112741.
Chen, X., S. Schauder, N. Potier, et al. 2002. “Structural Identification of a Bacterial Quorum-Sensing Signal Containing Boron.” Nature415, no. 6871: 545–549. https://doi.org/10.1038/415545a.
Cheng, Y., Y. Zhang, Z. Zhao, et al. 2022. “Guanidinium-Decorated Nanostructure for Precision Sonodynamic-Catalytic Therapy of MRSA-Infected Osteomyelitis.” Advanced Materials34, no. 50: e2206646. https://doi.org/10.1002/adma.202206646.
Chernin, L. S., M. K. Winson, J. M. Thompson, et al. 1998. “Chitinolytic Activity in Chromobacterium violaceum: Substrate Analysis and Regulation by Quorum Sensing.” Journal of Bacteriology180, no. 17: 4435–4441. https://doi.org/10.1128/JB.180.17.4435-4441.1998.
Chevrot, R., R. Rosen, E. Haudecoeur, et al. 2006. “GABA Controls the Level of Quorum-Sensing Signal in Agrobacterium tumefaciens.” Proceedings of the National Academy of Sciences103, no. 19: 7460–7464. https://doi.org/10.1073/pnas.0600313103.
Chilton, M. D., M. H. Drummond, D. J. Merlo, et al. 1977. “Stable Incorporation of Plasmid DNA into Higher Plant Cells: The Molecular Basis of Crown Gall Tumorigenesis.” Cell11, no. 2: 263–271. https://doi.org/10.1016/0092-8674(77)90043-5.
Chung, J., E. Goo, S. Yu, et al. 2011. “Small-Molecule Inhibitor Binding to an N-Acyl-Homoserine Lactone Synthase.” Proceedings of the National Academy of Sciences108, no. 29: 12089–12094. https://doi.org/10.1073/pnas.1103165108.
Cole, S. P., J. Harwood, R. Lee, R. She, and D. G. Guiney. 2004. “Characterization of Monospecies Biofilm Formation by Helicobacter pylori.” Journal of Bacteriology186, no. 10: 3124–3132. https://doi.org/10.1128/JB.186.10.3124-3132.2004.
Costerton, J. W., Z. Lewandowski, D. E. Caldwell, D. R. Korber, and H. M. Lappin-Scott. 1995. “Microbial Biofilms.” Annual Review of Microbiology49: 711–745. https://doi.org/10.1146/annurev.mi.49.100195.003431.
Cvitkovitch, D. G., Y. H. Li, and R. P. Ellen. 2003. “Quorum Sensing and Biofilm Formation in Streptococcal Infections.” Journal of Clinical Investigation112, no. 11: 1626–1632. https://doi.org/10.1172/JCI20430.
Dastgheyb, S. S., A. E. Villaruz, K. Y. Le, et al. 2015. “Role of Phenol-Soluble Modulins in Formation of Staphylococcus aureus Biofilms in Synovial Fluid.” Infection and Immunity83, no. 7: 2966–2975. https://doi.org/10.1128/IAI.00394-15.
Dawid, S., A. M. Roche, and J. N. Weiser. 2007. “The blp Bacteriocins of Streptococcus pneumoniae Mediate Intraspecies Competition Both In Vitro and In Vivo.” Infection and Immunity75, no. 1: 443–451. https://doi.org/10.1128/IAI.01775-05.
Debroas, D., A. Mone, and A. Ter Halle. 2017. “Plastics in the North Atlantic Garbage Patch: A Boat-Microbe for Hitchhikers and Plastic Degraders.” Science of the Total Environment599–600: 1222–1232. https://doi.org/10.1016/j.scitotenv.2017.05.059.
Defoirdt, T.2018. “Quorum-Sensing Systems as Targets for Antivirulence Therapy.” Trends in Microbiology26, no. 4: 313–328. https://doi.org/10.1016/j.tim.2017.10.005.
Defoirdt, T., N. Boon, P. Sorgeloos, W. Verstraete, and P. Bossier. 2008. “Quorum Sensing and Quorum Quenching in Vibrio harveyi: Lessons Learned From In Vivo Work.” ISME Journal2, no. 1: 19–26. https://doi.org/10.1038/ismej.2007.92.
Defoirdt, T., P. Bossier, P. Sorgeloos, and W. Verstraete. 2005. “The Impact of Mutations in the Quorum Sensing Systems of Aeromonas hydrophila, Streptococcus pneumoniae and Vibrio harveyi on Their Virulence Towards Gnotobiotically Cultured Artemia franciscana.” Environmental Microbiology7, no. 8: 1239–1247. https://doi.org/10.1111/j.1462-2920.2005.00807.x.
Defoirdt, T., C. M. Miyamoto, T. K. Wood, et al. 2007. “The Natural Furanone (5Z)-4-bromo-5-(bromomethylene)-3-butyl-2(5H)-furanone Disrupts Quorum Sensing-Regulated Gene Expression in Vibrio harveyi by Decreasing the DNA-Binding Activity of the Transcriptional Regulator Protein luxR.” Environmental Microbiology9, no. 10: 2486–2495. https://doi.org/10.1111/j.1462-2920.2007.01367.x.
Defoirdt, T., P. Sorgeloos, and P. Bossier. 2011. “Alternatives to Antibiotics for the Control of Bacterial Disease in Aquaculture.” Current Opinion in Microbiology14, no. 3: 251–258. https://doi.org/10.1016/j.mib.2011.03.004.
de Kievit, T. R., and B. H. Iglewski. 2000. “Bacterial Quorum Sensing in Pathogenic Relationships.” Infection and Immunity68, no. 9: 4839–4849. https://doi.org/10.1128/IAI.68.9.4839-4849.2000.
Deng, Z., X. M. Luo, J. Liu, and H. Wang. 2020. “Quorum Sensing, Biofilm, and Intestinal Mucosal Barrier: Involvement the Role of Probiotic.” Frontiers in Cellular and Infection Microbiology10: 538077. https://doi.org/10.3389/fcimb.2020.538077.
Diggle, S. P., K. Winzer, S. R. Chhabra, K. E. Worrall, M. Cámara, and P. Williams. 2003. “The Pseudomonas aeruginosa Quinolone Signal Molecule Overcomes the Cell Density-Dependency of the Quorum Sensing Hierarchy, Regulates rhl-Dependent Genes at the Onset of Stationary Phase and Can Be Produced in the Absence of LasR.” Molecular Microbiology50, no. 1: 29–43. https://doi.org/10.1046/j.1365-2958.2003.03672.x.
Domenech, A., A. R. Brochado, V. Sender, et al. 2020. “Proton Motive Force Disruptors Block Bacterial Competence and Horizontal Gene Transfer.” Cell Host & Microbe27, no. 4: 544–555.e3. https://doi.org/10.1016/j.chom.2020.02.002.
Dong, Y. H., J. L. Xu, X. Z. Li, and L. H. Zhang. 2000. “AiiA, an Enzyme That Inactivates the Acylhomoserine Lactone Quorum-Sensing Signal and Attenuates the Virulence of Erwinia carotovora.” Proceedings of the National Academy of Sciences97, no. 7: 3526–3531. https://doi.org/10.1073/pnas.97.7.3526.
Eberhard, A., A. L. Burlingame, C. Eberhard, G. L. Kenyon, K. H. Nealson, and N. J. Oppenheimer. 1981. “Structural Identification of Autoinducer of Photobacterium fischeri Luciferase.” Biochemistry20, no. 9: 2444–2449. https://doi.org/10.1021/bi00512a013.
Edel, M., H. Horn, and J. Gescher. 2019. “Biofilm Systems as Tools in Biotechnological Production.” Applied Microbiology and Biotechnology103, no. 13: 5095–5103. https://doi.org/10.1007/s00253-019-09869-x.
Engebrecht, J., K. Nealson, and M. Silverman. 1983. “Bacterial Bioluminescence: Isolation and Genetic Analysis of Functions From Vibrio fischeri.” Cell32, no. 3: 773–781. https://doi.org/10.1016/0092-8674(83)90063-6.
Engebrecht, J., and M. Silverman. 1984. “Identification of Genes and Gene Products Necessary for Bacterial Bioluminescence.” Proceedings of the National Academy of Sciences81, no. 13: 4154–4158. https://doi.org/10.1073/pnas.81.13.4154.
Engebrecht, J., and M. Silverman. 1987. “Nucleotide Sequence of the Regulatory Locus Controlling Expression of Bacterial Genes for Bioluminescence.” Nucleic Acids Research15, no. 24: 10455–10467. https://doi.org/10.1093/nar/15.24.10455.
Fan, Q., J. Zuo, H. Wang, D. Grenier, L. Yi, and Y. Wang. 2022. “Contribution of Quorum Sensing to Virulence and Antibiotic Resistance in Zoonotic Bacteria.” Biotechnology Advances59: 107965. https://doi.org/10.1016/j.biotechadv.2022.107965.
Federle, M. J., and B. L. Bassler. 2003. “Interspecies Communication in Bacteria.” Journal of Clinical Investigation112, no. 9: 1291–1299. https://doi.org/10.1172/JCI20195.
Forbes, N. S., R. S. Coffin, L. Deng, et al. 2018. “White Paper on Microbial Anti-Cancer Therapy and Prevention.” Journal for Immunotherapy of Cancer6, no. 1: 78. https://doi.org/10.1186/s40425-018-0381-3.
Forbes, N. S., L. L. Munn, D. Fukumura, and R. K. Jain. 2003. “Sparse Initial Entrapment of Systemically Injected Salmonella typhimurium Leads to Heterogeneous Accumulation Within Tumors.” Cancer Research63, no. 17: 5188–5193. https://www.ncbi.nlm.nih.gov/pubmed/14500342.
Freeman, J. A., and B. L. Bassler. 1999a. “A Genetic Analysis of the Function of LuxO, a Two-Component Response Regulator Involved in Quorum Sensing in Vibrio harveyi.” Molecular Microbiology31, no. 2: 665–677. https://doi.org/10.1046/j.1365-2958.1999.01208.x.
Freeman, J. A., and B. L. Bassler. 1999b. “Sequence and Function of LuxU: A Two-Component Phosphorelay Protein That Regulates Quorum Sensing in Vibrio harveyi.” Journal of Bacteriology181, no. 3: 899–906. https://doi.org/10.1128/JB.181.3.899-906.1999.
Freeman, J. A., B. N. Lilley, and B. L. Bassler. 2000. “A Genetic Analysis of the Functions of LuxN: A Two-Component Hybrid Sensor Kinase That Regulates Quorum Sensing in Vibrio harveyi.” Molecular Microbiology35, no. 1: 139–149. https://doi.org/10.1046/j.1365-2958.2000.01684.x.
Fueser, H., M. T. Mueller, and W. Traunspurger. 2020. “Ingestion of Microplastics by Meiobenthic Communities in Small-Scale Microcosm Experiments.” Science of the Total Environment746: 141276. https://doi.org/10.1016/j.scitotenv.2020.141276.
Fuqua, W. C., S. C. Winans, and E. P. Greenberg. 1994. “Quorum Sensing in Bacteria: The LuxR–LuxI Family of Cell Density-Responsive Transcriptional Regulators.” Journal of Bacteriology176, no. 2: 269–275. https://doi.org/10.1128/jb.176.2.269-275.1994.
Gallagher, L. A., S. L. McKnight, M. S. Kuznetsova, E. C. Pesci, and C. Manoil. 2002. “Functions Required for Extracellular Quinolone Signaling by Pseudomonas aeruginosa.” Journal of Bacteriology184, no. 23: 6472–6480. https://doi.org/10.1128/JB.184.23.6472-6480.2002.
Gambello, M. J., and B. H. Iglewski. 1991. “Cloning and Characterization of the Pseudomonas aeruginosa lasR Gene, a Transcriptional Activator of Elastase Expression.” Journal of Bacteriology173, no. 9: 3000–3009. https://doi.org/10.1128/jb.173.9.3000-3009.1991.
Ganai, S., R. B. Arenas, and N. S. Forbes. 2009. “Tumour-Targeted Delivery of TRAIL Using Salmonella typhimurium Enhances Breast Cancer Survival in Mice.” British Journal of Cancer101, no. 10: 1683–1691. https://doi.org/10.1038/sj.bjc.6605403.
Gao, M., H. Chen, A. Eberhard, et al. 2005. “sinI- and expR-Dependent Quorum Sensing in Sinorhizobium meliloti.” Journal of Bacteriology187, no. 23: 7931–7944. https://doi.org/10.1128/JB.187.23.7931-7944.2005.
Garmyn, D., L. Gal, J. P. Lemaître, A. Hartmann, and P. Piveteau. 2009. “Communication and Autoinduction in the Species Listeria monocytogenes: A Central Role for the agr System.” Communicative & Integrative Biology2, no. 4: 371–374. https://doi.org/10.4161/cib.2.4.8610.
Gómez-Gómez, B., L. Arregui, S. Serrano, A. Santos, T. Pérez-Corona, and Y. Madrid. 2019. “Selenium and Tellurium-Based Nanoparticles as Interfering Factors in Quorum Sensing-Regulated Processes: Violacein Production and Bacterial Biofilm Formation.” Metallomics11, no. 6: 1104–1114. https://doi.org/10.1039/c9mt00044e.
Graf, J., and E. G. Ruby. 1998. “Host-Derived Amino Acids Support the Proliferation of Symbiotic Bacteria.” Proceedings of the National Academy of Sciences95, no. 4: 1818–1822. https://doi.org/10.1073/pnas.95.4.1818.
Guan, Y., C. Y. Tsao, D. N. Quan, et al. 2018. “Focusing Quorum Sensing Signalling by Nano-Magnetic Assembly.” Environmental Microbiology20, no. 7: 2585–2597. https://doi.org/10.1111/1462-2920.14284.
Gurich, N., and J. E. González. 2009. “Role of Quorum Sensing in Sinorhizobium meliloti-Alfalfa Symbiosis.” Journal of Bacteriology191, no. 13: 4372–4382. https://doi.org/10.1128/JB.00376-09.
Hall, C. W., and T. F. Mah. 2017. “Molecular Mechanisms of Biofilm-Based Antibiotic Resistance and Tolerance in Pathogenic Bacteria.” FEMS Microbiology Reviews41, no. 3: 276–301. https://doi.org/10.1093/femsre/fux010.
Hall, S., C. McDermott, S. Anoopkumar-Dukie, et al. 2016. “Cellular Effects of Pyocyanin, a Secreted Virulence Factor of Pseudomonas aeruginosa.” Toxins8, no. 8: 236. https://doi.org/10.3390/toxins8080236.
Hamoen, L. W.2003. “Controlling Competence in Bacillus subtilis: Shared Use of Regulators.” Microbiology149, no. Pt 1: 9–17. https://doi.org/10.1099/mic.0.26003-0.
Han, X., and C. Lu. 2009. “Biological Activity and Identification of a Peptide Inhibitor of LuxS From Streptococcus suis Serotype 2.” FEMS Microbiology Letters294, no. 1: 16–23. https://doi.org/10.1111/j.1574-6968.2009.01534.x.
Haudecoeur, E., and D. Faure. 2010. “A Fine Control of Quorum-Sensing Communication in Agrobacterium tumefaciens.” Communicative & Integrative Biology3, no. 2: 84–88. https://doi.org/10.4161/cib.3.2.10429.
Hentzer, M., K. Riedel, T. B. Rasmussen, et al. 2002. “Inhibition of Quorum Sensing in Pseudomonas aeruginosa Biofilm Bacteria by a Halogenated Furanone Compound.” Microbiology148, no. Pt 1: 87–102. https://doi.org/10.1099/00221287-148-1-87.
Hirakawa, H., and H. Tomita. 2013. “Interference of Bacterial Cell-to-Cell Communication: A New Concept of Antimicrobial Chemotherapy Breaks Antibiotic.” Frontiers in Microbiology4: 114. https://doi.org/10.3389/fmicb.2013.00114.
Hornby, J. M., E. C. Jensen, A. D. Lisec, et al. 2001. “Quorum Sensing in the Dimorphic Fungus Candida albicans Is Mediated by Farnesol.” Applied and Environmental Microbiology67, no. 7: 2982–2992. https://doi.org/10.1128/AEM.67.7.2982-2992.2001.
Hu, J., S. Chen, Y. Yang, et al. 2022. “A Smart Hydrogel With Anti-Biofilm and Anti-Virulence Activities to Treat Pseudomonas aeruginosa Infections.” Advanced Healthcare Materials11, no. 13: e2200299. https://doi.org/10.1002/adhm.202200299.
Ivar do Sul, J. A., Â. Spengler, and M. F. Costa. 2009. “Here, There and Everywhere. Small Plastic Fragments and Pellets on Beaches of Fernando de Noronha (Equatorial Western Atlantic).” Marine Pollution Bulletin58, no. 8: 1236–1238. https://doi.org/10.1016/j.marpolbul.2009.05.004.
Jantaruk, P., W. Pabuprapap, A. Nakaew, D. Kunthalert, and A. Suksamrarn. 2021. “4-Methoxybenzalacetone, the Cinnamic Acid Analog as a Potential Quorum Sensing Inhibitor Against Chromobacterium violaceum and Pseudomonas aeruginosa.” World Journal of Microbiology and Biotechnology37, no. 9: 153. https://doi.org/10.1007/s11274-021-03119-x.
Jin, Y., W. Li, H. Zhang, X. Ba, Z. Li, and J. Zhou. 2023. “The Post-Antibiotic Era: A New Dawn for Bacteriophages.” Biology12, no. 5: 681. https://doi.org/10.3390/biology12050681.
Kalia, V. C., S. K. S. Patel, B. K. Cho, T. K. Wood, and J. K. Lee. 2022. “Emerging Applications of Bacteria as Antitumor Agents.” Seminars in Cancer Biology86, no. Pt 2: 1014–1025. https://doi.org/10.1016/j.semcancer.2021.05.012.
Kalia, V. C., S. K. S. Patel, Y. C. Kang, and J. K. Lee. 2019. “Quorum Sensing Inhibitors as Antipathogens: Biotechnological Applications.” Biotechnology Advances37, no. 1: 68–90. https://doi.org/10.1016/j.biotechadv.2018.11.006.
Kalia, V. C., A. Rani, S. Lal, S. Cheema, and C. P. Raut. 2007. “Combing Databases Reveals Potential Antibiotic Producers.” Expert Opinion on Drug Discovery2, no. 2: 211–224. https://doi.org/10.1517/17460441.2.2.211.
Kamli, M. R., M. A. Malik, V. Srivastava, J. S. M. Sabir, E. H. Mattar, and A. Ahmad. 2021. “Biogenic ZnO Nanoparticles Synthesized From Origanum vulgare Abrogates Quorum Sensing and Biofilm Formation in Opportunistic Pathogen Chromobacterium violaceum.” Pharmaceutics13, no. 11: 1743. https://doi.org/10.3390/pharmaceutics13111743.
Karlsson, D., S. Karlsson, E. Gustafsson, B. H. Normark, and P. Nilsson. 2007. “Modeling the Regulation of the Competence-Evoking Quorum Sensing Network in Streptococcus pneumoniae.” Biosystems90, no. 1: 211–223. https://doi.org/10.1016/j.biosystems.2006.08.005.
Khaleghi, M., F. Haghi, M. Gholami, et al. 2023. “A Fabricated Hydrogel of Hyaluronic Acid/Curcumin Shows Super-Activity to Heal the Bacterial Infected Wound.” AMB Express13, no. 1: 29. https://doi.org/10.1186/s13568-023-01533-y.
Khan, F., A. Javaid, and Y. M. Kim. 2019. “Functional Diversity of Quorum Sensing Receptors in Pathogenic Bacteria: Interspecies, Intraspecies and Interkingdom Level.” Current Drug Targets20, no. 6: 655–667. https://doi.org/10.2174/1389450120666181123123333.
Knutsen, E., O. Ween, and L. S. Håvarstein. 2004. “Two Separate Quorum-Sensing Systems Upregulate Transcription of the Same ABC Transporter in Streptococcus pneumoniae.” Journal of Bacteriology186, no. 10: 3078–3085. https://doi.org/10.1128/JB.186.10.3078-3085.2004.
Koutsoudis, M. D., D. Tsaltas, T. D. Minogue, and S. B. von Bodman. 2006. “Quorum-Sensing Regulation Governs Bacterial Adhesion, Biofilm Development, and Host Colonization in Pantoea stewartii Subspecies stewartii.” Proceedings of the National Academy of Sciences103, no. 15: 5983–5988. https://doi.org/10.1073/pnas.0509860103.
Kuang, Z., Y. Hao, B. E. Walling, J. L. Jeffries, D. E. Ohman, and G. W. Lau. 2011. “Pseudomonas aeruginosa Elastase Provides an Escape From Phagocytosis by Degrading the Pulmonary Surfactant Protein-A.” PLoS ONE6, no. 11: e27091. https://doi.org/10.1371/journal.pone.0027091.
Kumar, L., S. K. S. Patel, K. Kharga, et al. 2022. “Molecular Mechanisms and Applications of N-Acyl Homoserine Lactone-Mediated Quorum Sensing in Bacteria.” Molecules27, no. 21: 7584. https://doi.org/10.3390/molecules27217584.
Kuo, A., S. M. Callahan, and P. V. Dunlap. 1996. “Modulation of Luminescence Operon Expression by N-Octanoyl-l-Homoserine Lactone in ainS Mutants of Vibrio fischeri.” Journal of Bacteriology178, no. 4: 971–976. https://doi.org/10.1128/jb.178.4.971-976.1996.
Landman, C., J. P. Grill, J. M. Mallet, et al. 2018. “Inter-Kingdom Effect on Epithelial Cells of the N-Acyl Homoserine Lactone 3-oxo-C12:2, a Major Quorum-Sensing Molecule From Gut Microbiota.” PLoS ONE13, no. 8: e0202587. https://doi.org/10.1371/journal.pone.0202587.
Lang, J., and D. Faure. 2014. “Functions and Regulation of Quorum-Sensing in Agrobacterium tumefaciens.” Frontiers in Plant Science5: 14. https://doi.org/10.3389/fpls.2014.00014.
Latifi, A., M. Foglino, K. Tanaka, P. Williams, and A. Lazdunski. 1996. “A Hierarchical Quorum-Sensing Cascade in Pseudomonas aeruginosa Links the Transcriptional Activators LasR and RhIR (VsmR) to Expression of the Stationary-Phase Sigma Factor RpoS.” Molecular Microbiology21, no. 6: 1137–1146. https://doi.org/10.1046/j.1365-2958.1996.00063.x.
Lee, J., and L. Zhang. 2015. “The Hierarchy Quorum Sensing Network in Pseudomonas aeruginosa.” Protein & Cell6, no. 1: 26–41. https://doi.org/10.1007/s13238-014-0100-x.
Lenz, D. H., K. C. Mok, B. N. Lilley, R. V. Kulkarni, N. S. Wingreen, and B. L. Bassler. 2004. “The Small RNA Chaperone Hfq and Multiple Small RNAs Control Quorum Sensing in Vibrio harveyi and Vibrio cholerae.” Cell118, no. 1: 69–82. https://doi.org/10.1016/j.cell.2004.06.009.
Lewenza, S., B. Conway, E. P. Greenberg, and P. A. Sokol. 1999. “Quorum Sensing in Burkholderia cepacia: Identification of the LuxRI Homologs CepRI.” Journal of Bacteriology181, no. 3: 748–756. https://doi.org/10.1128/JB.181.3.748-756.1999.
Li, J., and X. Zhao. 2020. “Effects of Quorum Sensing on the Biofilm Formation and Viable But Non-Culturable State.” Food Research International137: 109742. https://doi.org/10.1016/j.foodres.2020.109742.
Liang, X., Q. Y. Chen, G. M. Seabra, et al. 2021. “Bifunctional Doscadenamides Activate Quorum Sensing in Gram-Negative Bacteria and Synergize With TRAIL to Induce Apoptosis in Cancer Cells.” Journal of Natural Products84, no. 3: 779–789. https://doi.org/10.1021/acs.jnatprod.0c01003.
Lilley, B. N., and B. L. Bassler. 2000. “Regulation of Quorum Sensing in Vibrio harveyi by LuxO and Sigma-54.” Molecular Microbiology36, no. 4: 940–954. https://doi.org/10.1046/j.1365-2958.2000.01913.x.
Lin, Y. H., J. L. Xu, J. Hu, et al. 2003. “Acyl-Homoserine Lactone Acylase From Ralstonia Strain XJ12B Represents a Novel and Potent Class of Quorum-Quenching Enzymes.” Molecular Microbiology47, no. 3: 849–860. https://doi.org/10.1046/j.1365-2958.2003.03351.x.
Loessner, H., A. Endmann, S. Leschner, et al. 2007. “Remote Control of Tumour-Targeted Salmonella enterica Serovar Typhimurium by the Use of l-Arabinose as Inducer of Bacterial Gene Expression In Vivo.” Cellular Microbiology9, no. 6: 1529–1537. https://doi.org/10.1111/j.1462-5822.2007.00890.x.
Lu, J., S. Zhang, S. Gao, P. Wang, P. L. Bond, and J. Guo. 2020. “New Insights of the Bacterial Response to Exposure of Differently Sized Silver Nanomaterials.” Water Research169: 115205. https://doi.org/10.1016/j.watres.2019.115205.
Luong, T., A. C. Salabarria, and D. R. Roach. 2020. “Phage Therapy in the Resistance Era: Where Do We Stand and Where Are We Going?” Clinical Therapeutics42, no. 9: 1659–1680. https://doi.org/10.1016/j.clinthera.2020.07.014.
Maddela, N. R., B. Sheng, S. Yuan, Z. Zhou, R. Villamar-Torres, and F. Meng. 2019. “Roles of Quorum Sensing in Biological Wastewater Treatment: A Critical Review.” Chemosphere221: 616–629. https://doi.org/10.1016/j.chemosphere.2019.01.064.
Maddula, V. S. R. K., Z. Zhang, E. A. Pierson, and L. S. Pierson, 3rd. 2006. “Quorum Sensing and Phenazines Are Involved in Biofilm Formation by Pseudomonas chlororaphis (aureofaciens) Strain 30–84.” Microbial Ecology52, no. 2: 289–301. https://doi.org/10.1007/s00248-006-9064-6.
Manefield, M., L. Harris, S. A. Rice, R. de Nys, and S. Kjelleberg. 2000. “Inhibition of Luminescence and Virulence in the Black Tiger Prawn (Penaeus monodon) Pathogen Vibrio harveyi by Intercellular Signal Antagonists.” Applied and Environmental Microbiology66, no. 5: 2079–2084. https://doi.org/10.1128/AEM.66.5.2079-2084.2000.
Matsumoto, M., S. Nakagawa, L. Zhang, et al. 2021. “Interaction Between Staphylococcus Agr Virulence and Neutrophils Regulates Pathogen Expansion in the Skin.” Cell Host & Microbe29, no. 6: 930–940.e4. https://doi.org/10.1016/j.chom.2021.03.007.
Mauritzen, J. J., E. Søndberg, P. G. Kalatzis, et al. 2023. “Strain-Specific Quorum-Sensing Responses Determine Virulence Properties in Vibrio anguillarum.” Environmental Microbiology25, no. 7: 1344–1362. https://doi.org/10.1111/1462-2920.16356.
Mayer, C., M. Romero, A. Muras, and A. Otero. 2015. “Aii20J, a Wide-Spectrum Thermostable N-Acylhomoserine Lactonase From the Marine Bacterium Tenacibaculum sp. 20J, Can Quench AHL-Mediated Acid Resistance in Escherichia coli.” Applied Microbiology and Biotechnology99, no. 22: 9523–9539. https://doi.org/10.1007/s00253-015-6741-8.
Mayville, P., G. Ji, R. Beavis, et al. 1999. “Structure-Activity Analysis of Synthetic Autoinducing Thiolactone Peptides From Staphylococcus aureus Responsible for Virulence.” Proceedings of the National Academy of Sciences96, no. 4: 1218–1223. https://doi.org/10.1073/pnas.96.4.1218.
Mazzola, M., R. J. Cook, L. S. Thomashow, D. M. Weller, and L. S. Pierson, 3rd. 1992. “Contribution of Phenazine Antibiotic Biosynthesis to the Ecological Competence of Fluorescent Pseudomonads in Soil Habitats.” Applied and Environmental Microbiology58, no. 8: 2616–2624. https://doi.org/10.1128/aem.58.8.2616-2624.1992.
Merritt, J., F. Qi, S. D. Goodman, M. H. Anderson, and W. Shi. 2003. “Mutation of Luxs Affects Biofilm Formation in Streptococcus mutans.” Infection and Immunity71, no. 4: 1972–1979. https://doi.org/10.1128/IAI.71.4.1972-1979.2003.
Miller, C., and J. Gilmore. 2020. “Detection of Quorum-Sensing Molecules for Pathogenic Molecules Using Cell-Based and Cell-Free Biosensors.” Antibiotics (USSR)9, no. 5: 259. https://doi.org/10.3390/antibiotics9050259.
Miquel, S., R. Lagrafeuille, B. Souweine, and C. Forestier. 2016. “Anti-Biofilm Activity as a Health Issue.” Frontiers in Microbiology7: 592. https://doi.org/10.3389/fmicb.2016.00592.
Miyamoto, C. M., Y. H. Lin, and E. A. Meighen. 2000. “Control of Bioluminescence in Vibrio fischeri by the LuxO Signal Response Regulator.” Molecular Microbiology36, no. 3: 594–607. https://doi.org/10.1046/j.1365-2958.2000.01875.x.
Mogosanu, G., A. Grumezescu, K. S. Huang, L. Bejenaru, and C. Bejenaru. 2015. “Prevention of Microbial Communities: Novel Approaches Based Natural Products.” Current Pharmaceutical Biotechnology16, no. 2: 94–111. https://doi.org/10.2174/138920101602150112145916.
Moré, M. I., L. D. Finger, J. L. Stryker, C. Fuqua, A. Eberhard, and S. C. Winans. 1996. “Enzymatic Synthesis of a Quorum-Sensing Autoinducer Through Use of Defined Substrates.” Science272, no. 5268: 1655–1658. https://doi.org/10.1126/science.272.5268.1655.
Morrison, J.2012. “The Staphylococcal Accessory Regulator, SarA, Is an RNA-Binding Protein That Modulates the mRNA Turnover Properties of Late-Exponential and Stationary Phase Staphylococcus aureus Cells.” Frontiers in Cellular and Infection Microbiology2: 26. https://doi.org/10.3389/fcimb.2012.00026.
Mukherjee, S., D. Moustafa, C. D. Smith, J. B. Goldberg, and B. L. Bassler. 2017. “The RhlR Quorum-Sensing Receptor Controls Pseudomonas aeruginosa Pathogenesis and Biofilm Development Independently of Its Canonical Homoserine Lactone Autoinducer.” PLoS Pathogens13, no. 7: e1006504. https://doi.org/10.1371/journal.ppat.1006504.
Nasser, W., M. L. Bouillant, G. Salmond, and S. Reverchon. 1998. “Characterization of the Erwinia chrysanthemi expI–expR locus directing the Synthesis of Two N-Acyl-Homoserine Lactone Signal Molecules.” Molecular Microbiology29, no. 6: 1391–1405. https://doi.org/10.1046/j.1365-2958.1998.01022.x.
Nealson, K. H., T. Platt, and J. W. Hastings. 1970. “Cellular Control of the Synthesis and Activity of the Bacterial Luminescent System.” Journal of Bacteriology104, no. 1: 313–322. https://doi.org/10.1128/jb.104.1.313-322.1970.
Neiditch, M. B., G. C. Capodagli, G. Prehna, and M. J. Federle. 2017. “Genetic and Structural Analyses of RRNPP Intercellular Peptide Signaling of Gram-Positive Bacteria.” Annual Review of Genetics51: 311–333. https://doi.org/10.1146/annurev-genet-120116-023507.
Ng, Y. K., M. Grasso, V. Wright, V. Garcia, P. Williams, and S. Atkinson. 2018. “The Quorum Sensing System of Yersinia enterocolitica 8081 Regulates Swimming Motility, Host Cell Attachment, and Virulence Plasmid Maintenance.” Genes9, no. 6: 307. https://doi.org/10.3390/genes9060307.
Novick, R. P.2003. “Autoinduction and Signal Transduction in the Regulation of Staphylococcal Virulence.” Molecular Microbiology48, no. 6: 1429–1449. https://doi.org/10.1046/j.1365-2958.2003.03526.x.
Novick, R. P., H. F. Ross, S. J. Projan, J. Kornblum, B. Kreiswirth, and S. Moghazeh. 1993. “Synthesis of Staphylococcal Virulence Factors Is Controlled by a Regulatory RNA Molecule.” EMBO Journal12, no. 10: 3967–3975. https://doi.org/10.1002/j.1460-2075.1993.tb06074.x.
Ochsner, U. A., A. K. Koch, A. Fiechter, and J. Reiser. 1994. “Isolation and Characterization of a Regulatory Gene Affecting Rhamnolipid Biosurfactant Synthesis in Pseudomonas aeruginosa.” Journal of Bacteriology176, no. 7: 2044–2054. https://doi.org/10.1128/jb.176.7.2044-2054.1994.
O'Loughlin, C. T., L. C. Miller, A. Siryaporn, K. Drescher, M. F. Semmelhack, and B. L. Bassler. 2013. “A Quorum-Sensing Inhibitor Blocks Pseudomonas aeruginosa Virulence and Biofilm Formation.” Proceedings of the National Academy of Sciences110, no. 44: 17981–17986. https://doi.org/10.1073/pnas.1316981110.
Omwenga, E. O., A. Hensel, A. Shitandi, and F. M. Goycoolea. 2018. “Chitosan Nanoencapsulation of Flavonoids Enhances Their Quorum Sensing and Biofilm Formation Inhibitory Activities Against an E. coli Top 10 Biosensor.” Colloids and Surfaces B: Biointerfaces164: 125–133. https://doi.org/10.1016/j.colsurfb.2018.01.019.
Packiavathy, I. A. S. V., A. Kannappan, S. Thiyagarajan, et al. 2021. “AHL-Lactonase Producing Psychrobacter sp. From Palk Bay Sediment Mitigates Quorum Sensing-Mediated Virulence Production in Gram Negative Bacterial Pathogens.” Frontiers in Microbiology12: 634593. https://doi.org/10.3389/fmicb.2021.634593.
Paluch, E., A. Piecuch, E. Obłąk, Ł. Lamch, and K. A. Wilk. 2018. “Antifungal Activity of Newly Synthesized Chemodegradable Dicephalic-Type Cationic Surfactants.” Colloids and Surfaces B: Biointerfaces164: 34–41. https://doi.org/10.1016/j.colsurfb.2018.01.020.
Paluch, E., J. Rewak-Soroczyńska, I. Jędrusik, E. Mazurkiewicz, and K. Jermakow. 2020. “Prevention of Biofilm Formation by Quorum Quenching.” Applied Microbiology and Biotechnology104, no. 5: 1871–1881. https://doi.org/10.1007/s00253-020-10349-w.
Parga, A., A. Muras, P. Otero-Casal, et al. 2023. “The Quorum Quenching Enzyme Aii20J Modifies In Vitro Periodontal Biofilm Formation.” Frontiers in Cellular and Infection Microbiology13: 1118630. https://doi.org/10.3389/fcimb.2023.1118630.
Passador, L., J. M. Cook, M. J. Gambello, L. Rust, and B. H. Iglewski. 1993. “Expression of Pseudomonas aeruginosa Virulence Genes Requires Cell-to-Cell Communication.” Science260, no. 5111: 1127–1130. https://doi.org/10.1126/science.8493556.
Passos da Silva, D., M. Schofield, M. Parsek, and B. Tseng. 2017. “An Update on the Sociomicrobiology of Quorum Sensing in Gram-Negative Biofilm Development.” Pathogens6, no. 4: 51. https://doi.org/10.3390/pathogens6040051.
Patel, F., and S. D. Spassieva. 2018. “Side Effects in Cancer Therapy: Are Sphingolipids to Blame?” Advances in Cancer Research140: 367–388. https://doi.org/10.1016/bs.acr.2018.04.017.
Pearson, J. P., K. M. Gray, L. Passador, et al. 1994. “Structure of the Autoinducer Required for Expression of Pseudomonas aeruginosa Virulence Genes.” Proceedings of the National Academy of Sciences91, no. 1: 197–201. https://doi.org/10.1073/pnas.91.1.197.
Pearson, J. P., E. C. Pesci, and B. H. Iglewski. 1997. “Roles of Pseudomonas aeruginosa las and rhl Quorum-Sensing Systems in Control of Elastase and Rhamnolipid Biosynthesis Genes.” Journal of Bacteriology179, no. 18: 5756–5767. https://doi.org/10.1128/jb.179.18.5756-5767.1997.
Pearson, J. P., L. Passador, B. H. Iglewski, and E. P. Greenberg. 1995. “A Second N-Acylhomoserine Lactone Signal Produced by Pseudomonas aeruginosa.” Proceedings of the National Academy of Sciences92, no. 5: 1490–1494. https://doi.org/10.1073/pnas.92.5.1490.
Peng, X., J. Chen, Y. Gan, et al. 2024. “Biofunctional Lipid Nanoparticles for Precision Treatment and Prophylaxis of Bacterial Infections.” Science Advances10, no. 14: eadk9754. https://doi.org/10.1126/sciadv.adk9754.
Pesci, E. C., J. B. J. Milbank, J. P. Pearson, et al. 1999. “Quinolone Signaling in the Cell-to-Cell Communication System of Pseudomonas aeruginosa.” Proceedings of the National Academy of Sciences96, no. 20: 11229–11234. https://doi.org/10.1073/pnas.96.20.11229.
Pesci, E. C., J. P. Pearson, P. C. Seed, and B. H. Iglewski. 1997. “Regulation of las and rhl Quorum Sensing in Pseudomonas aeruginosa.” Journal of Bacteriology179, no. 10: 3127–3132. https://doi.org/10.1128/jb.179.10.3127-3132.1997.
Piecuch, A., Ł. Lamch, E. Paluch, E. Obłąk, and K. A. Wilk. 2016. “Biofilm Prevention by Dicephalic Cationic Surfactants and Their Interactions With DNA.” Journal of Applied Microbiology121, no. 3: 682–692. https://doi.org/10.1111/jam.13204.
Piletska, E. V., G. Stavroulakis, K. Karim, et al. 2010. “Attenuation of Vibrio fischeri Quorum Sensing Using Rationally Designed Polymers.” Biomacromolecules11, no. 4: 975–980. https://doi.org/10.1021/bm901451j.
Pragman, A. A., J. M. Yarwood, T. J. Tripp, and P. M. Schlievert. 2004. “Characterization of Virulence Factor Regulation by SrrAB, a Two-Component System in Staphylococcus aureus.” Journal of Bacteriology186, no. 8: 2430–2438. https://doi.org/10.1128/JB.186.8.2430-2438.2004.
Prateeksha, S. K. Barik, and B. N. Singh. 2019. “Nanoemulsion-Loaded Hydrogel Coatings for Inhibition of Bacterial Virulence and Biofilm Formation on Solid Surfaces.” Scientific Reports9, no. 1: 6520. https://doi.org/10.1038/s41598-019-43016-w.
Proctor, C. R., P. A. McCarron, and N. G. Ternan. 2020. “Furanone Quorum-Sensing Inhibitors With Potential as Novel Therapeutics Against Pseudomonas aeruginosa.” Journal of Medical Microbiology69, no. 2: 195–206. https://doi.org/10.1099/jmm.0.001144.
Pumbwe, L., C. A. Skilbeck, and H. M. Wexler. 2008. “Presence of Quorum-Sensing Systems Associated With Multidrug Resistance and Biofilm Formation in Bacteroides fragilis.” Microbial Ecology56, no. 3: 412–419. https://doi.org/10.1007/s00248-007-9358-3.
Qu, Y., Y. Zou, G. Wang, Y. Zhang, and Q. Yu. 2024. “Disruption of Communication: Recent Advances in Antibiofilm Materials With Anti-Quorum Sensing Properties.” ACS Applied Materials & Interfaces16, no. 11: 13353–13383. https://doi.org/10.1021/acsami.4c01428.
Ramić, D., A. Klančnik, S. S. Možina, and I. Dogsa. 2022. “Elucidation of the AI-2 Communication System in the Food-Borne Pathogen Campylobacter jejuni by Whole-Cell-Based Biosensor Quantification.” Biosensors and Bioelectronics212: 114439. https://doi.org/10.1016/j.bios.2022.114439.
Rehman, Z. U., and T. Leiknes. 2018. “Quorum-Quenching Bacteria Isolated From Red Sea Sediments Reduce Biofilm Formation by Pseudomonas aeruginosa.” Frontiers in Microbiology9: 1354. https://doi.org/10.3389/fmicb.2018.01354.
Reichmann, P., and R. Hakenbeck. 2000. “Allelic Variation in a Peptide-Inducible Two-Component System of Streptococcus pneumoniae.” FEMS Microbiology Letters190, no. 2: 231–236. https://doi.org/10.1111/j.1574-6968.2000.tb09291.x.
Reina, J. C., M. Romero, R. Salto, M. Cámara, and I. Llamas. 2021. “AhaP, A Quorum Quenching Acylase From Psychrobacter sp. M9-54-1 That Attenuates Pseudomonas aeruginosa and Vibrio coralliilyticus Virulence.” Marine Drugs19, no. 1: 16. https://doi.org/10.3390/md19010016.
Remuzgo-Martínez, S., M. Lázaro-Díez, C. Mayer, et al. 2015. “Biofilm Formation and Quorum-Sensing-Molecule Production by Clinical Isolates of Serratia liquefaciens.” Applied and Environmental Microbiology81, no. 10: 3306–3315. https://doi.org/10.1128/AEM.00088-15.
Reynolds, D., and M. Kollef. 2021. “The Epidemiology and Pathogenesis and Treatment of Pseudomonas aeruginosa Infections: An Update.” Drugs81, no. 18: 2117–2131. https://doi.org/10.1007/s40265-021-01635-6.
Reynolds, J., and S. Wigneshweraraj. 2011. “Molecular Insights into the Control of Transcription Initiation at the Staphylococcus aureus agr Operon.” Journal of Molecular Biology412, no. 5: 862–881. https://doi.org/10.1016/j.jmb.2011.06.018.
Ritzert, J. T., G. Minasov, R. Embry, M. J. Schipma, and K. J. F. Satchell. 2019. “The Cyclic AMP Receptor Protein Regulates Quorum Sensing and Global Gene Expression in Yersinia pestis During Planktonic Growth and Growth in Biofilms.” mBio10, no. 6: e02613-19. https://doi.org/10.1128/mBio.02613-19.
Roca, A., M. Cabeo, C. Enguidanos, F. Martínez-Checa, I. Sampedro, and I. Llamas. 2024. “Potential of the Quorum-Quenching and Plant-Growth Promoting Halotolerant Bacillus toyonensis AA1EC1 as Biocontrol Agent.” Microbial Biotechnology17, no. 3: e14420. https://doi.org/10.1111/1751-7915.14420.
Rogers, G. B., M. P. Carroll, and K. D. Bruce. 2012. “Enhancing the Utility of Existing Antibiotics by Targeting Bacterial Behaviour?” British Journal of Pharmacology165, no. 4: 845–857. https://doi.org/10.1111/j.1476-5381.2011.01643.x.
Rosemeyer, V., J. Michiels, C. Verreth, and J. Vanderleyden. 1998. “luxI- and luxR-Homologous Genes of Rhizobium etli CNPAF512 Contribute to Synthesis of Autoinducer Molecules and Nodulation of Phaseolus vulgaris.” Journal of Bacteriology180, no. 4: 815–821. https://doi.org/10.1128/JB.180.4.815-821.1998.
Rowe, S. E., V. Mahon, S. G. Smith, and J. P. O'Gara. 2011. “A Novel Role for SarX in Staphylococcus epidermidis Biofilm Regulation.” Microbiology157, no. Pt 4: 1042–1049. https://doi.org/10.1099/mic.0.046581-0.
Roy, S., G. Chowdhury, A. K. Mukhopadhyay, S. Dutta, and S. Basu. 2022. “Convergence of Biofilm Formation and Antibiotic Resistance in Acinetobacter baumannii Infection.” Frontiers in Medicine9: 793615. https://doi.org/10.3389/fmed.2022.793615.
Rutherford, S. T., J. C. van Kessel, Y. Shao, and B. L. Bassler. 2011. “AphA and LuxR/HapR Reciprocally Control Quorum Sensing in Vibrios.” Genes & Development25, no. 4: 397–408. https://doi.org/10.1101/gad.2015011.
Schuster, M., and E. Peter Greenberg. 2006. “A Network of Networks: Quorum-Sensing Gene Regulation in Pseudomonas aeruginosa.” International Journal of Medical Microbiology296, no. 2–3: 73–81. https://doi.org/10.1016/j.ijmm.2006.01.036.
Schwenteit, J., L. Gram, K. F. Nielsen, et al. 2011. “Quorum Sensing in Aeromonas salmonicida subsp. Achromogenes and the Effect of the Autoinducer Synthase Asai on Bacterial Virulence.” Veterinary Microbiology147, no. 3–4: 389–397. https://doi.org/10.1016/j.vetmic.2010.07.020.
Shah, S., S. Gaikwad, S. Nagar, et al. 2019. “Biofilm Inhibition and Anti-Quorum Sensing Activity of Phytosynthesized Silver Nanoparticles Against the Nosocomial Pathogen Pseudomonas aeruginosa.” Biofouling35, no. 1: 34–49. https://doi.org/10.1080/08927014.2018.1563686.
Shanker, E., and M. Federle. 2017. “Quorum Sensing Regulation of Competence and Bacteriocins in Streptococcus pneumoniae and mutans.” Genes8, no. 1: 15. https://doi.org/10.3390/genes8010015.
Sheikh, H., A. John, N. Musa, L. A. Abdulrazzak, M. Alfatama, and A. Fadhlina. 2022. “Vibrio spp. and Their Vibriocin as a Vibriosis Control Measure in Aquaculture.” Applied Biochemistry and Biotechnology194, no. 10: 4477–4491. https://doi.org/10.1007/s12010-022-03919-3.
Showalter, R. E., M. O. Martin, and M. R. Silverman. 1990. “Cloning and Nucleotide Sequence of luxR, a Regulatory Gene Controlling Bioluminescence in Vibrio harveyi.” Journal of Bacteriology172, no. 6: 2946–2954. https://doi.org/10.1128/jb.172.6.2946-2954.1990.
Silpe, J. E., and B. L. Bassler. 2019. “A Host-Produced Quorum-Sensing Autoinducer Controls a Phage Lysis-Lysogeny Decision.” Cell176, no. 1–2: 268–280.e13. https://doi.org/10.1016/j.cell.2018.10.059.
Silpe, J. E., O. P. Duddy, G. E. Johnson, et al. 2023. “Small Protein Modules Dictate Prophage Fates During Polylysogeny.” Nature620, no. 7974: 625–633. https://doi.org/10.1038/s41586-023-06376-y.
Skindersoe, M. E., P. Ettinger-Epstein, T. B. Rasmussen, T. Bjarnsholt, R. de Nys, and M. Givskov. 2008. “Quorum Sensing Antagonism From Marine Organisms.” Marine Biotechnology10, no. 1: 56–63. https://doi.org/10.1007/s10126-007-9036-y.
Solano, C., M. Echeverz, and I. Lasa. 2014. “Biofilm Dispersion and Quorum Sensing.” Current Opinion in Microbiology18: 96–104. https://doi.org/10.1016/j.mib.2014.02.008.
Steinmoen, H., E. Knutsen, and L. S. Håvarstein. 2002. “Induction of Natural Competence in Streptococcus pneumoniae Triggers Lysis and DNA Release From a Subfraction of the Cell Population.” Proceedings of the National Academy of Sciences99, no. 11: 7681–7686. https://doi.org/10.1073/pnas.112464599.
Stevenson, B., and K. Babb. 2002. “LuxS-Mediated Quorum Sensing in Borrelia burgdorferi, the Lyme Disease Spirochete.” Infection and Immunity70, no. 8: 4099–4105. https://doi.org/10.1128/IAI.70.8.4099-4105.2002.
Storz, M. P., G. Allegretta, B. Kirsch, M. Empting, and R. W. Hartmann. 2014. “From In Vitro to in Cellulo: Structure-Activity Relationship of (2-nitrophenyl)methanol Derivatives as Inhibitors of PqsD in Pseudomonas aeruginosa.” Organic and Biomolecular Chemistry12, no. 32: 6094–6104. https://doi.org/10.1039/c4ob00707g.
Su, Y., K. Tang, J. Liu, Y. Wang, Y. Zheng, and X. H. Zhang. 2018. “Quorum Sensing System of Ruegeria mobilis Rm01 Controls Lipase and Biofilm Formation.” Frontiers in Microbiology9: 3304. https://doi.org/10.3389/fmicb.2018.03304.
Sully, E. K., N. Malachowa, B. O. Elmore, et al. 2014. “Selective Chemical Inhibition of agr Quorum Sensing in Staphylococcus aureus Promotes Host Defense With Minimal Impact on Resistance.” PLoS Pathogens10, no. 6: e1004174. https://doi.org/10.1371/journal.ppat.1004174.
Sun, X., P. Hill, J. Liu, J. Qian, Y. Ma, and S. Zhou. 2021. “Marine-Source Quorum Quenching Enzyme YtnP to Improve Hygiene Quality in Dental Units.” Marine Drugs19, no. 4: 225. https://doi.org/10.3390/md19040225.
Sun, B., H. Luo, H. Jiang, Z. Wang, and A. Jia. 2021. “Inhibition of Quorum Sensing and Biofilm Formation of Esculetin on Aeromonas hydrophila.” Frontiers in Microbiology12: 737626. https://doi.org/10.3389/fmicb.2021.737626.
Suramo, I., M. Päivänsalo, A. Leinonen, and M. Pamilo. 1981. “Sonographische Darstellung von Hypernephromen.” RöFo - Fortschritte auf dem Gebiet der Röntgenstrahlen und der bildgebenden Verfahren135, no. 6: 649–652. https://doi.org/10.1055/s-2008-1056489.
Thompson, R. C., Y. Olsen, R. P. Mitchell, et al. 2004. “Lost at Sea: Where Is All the Plastic?” Science304, no. 5672: 838. https://doi.org/10.1126/science.1094559.
Tielker, D., S. Hacker, R. Loris, et al. 2005. “Pseudomonas aeruginosa Lectin LecB Is Located in the Outer Membrane and Is Involved in Biofilm Formation.” Microbiology151, no. Pt 5: 1313–1323. https://doi.org/10.1099/mic.0.27701-0.
Tomasz, A.1965. “Control of the Competent State in Pneumococcus by a Hormone-Like Cell Product: An Example for a New Type of Regulatory Mechanism in Bacteria.” Nature208, no. 5006: 155–159. https://doi.org/10.1038/208155a0.
Tu, C., H. Lu, T. Zhou, et al. 2022. “Promoting the Healing of Infected Diabetic Wound by an Anti-Bacterial and Nano-Enzyme-Containing Hydrogel With Inflammation-Suppressing, ROS-Scavenging, Oxygen and Nitric Oxide-Generating Properties.” Biomaterials286: 121597. https://doi.org/10.1016/j.biomaterials.2022.121597.
Turner, N. A., B. K. Sharma-Kuinkel, S. A. Maskarinec, et al. 2019. “Methicillin-Resistant Staphylococcus aureus: An Overview of Basic and Clinical Research.” Nature Reviews Microbiology17, no. 4: 203–218. https://doi.org/10.1038/s41579-018-0147-4.
Uroz, S., P. M. Oger, E. Chapelle, M. Adeline, D. Faure, and Y. Dessaux. 2008. “A Rhodococcus qsdA-Encoded Enzyme Defines a Novel Class of Large-Spectrum Quorum-Quenching Lactonases.” Applied and Environmental Microbiology74, no. 5: 1357–1366. https://doi.org/10.1128/AEM.02014-07.
Vasudevan, S., P. Srinivasan, P. Neelakantan, J. B. B. Rayappan, and A. P. Solomon. 2021. “Photoluminescence-Based Bioassay With Cysteamine-Capped TiO2 Nanoparticles for the Selective Recognition of N-Acyl Homoserine Lactones.” Frontiers in Bioengineering and Biotechnology9: 750933. https://doi.org/10.3389/fbioe.2021.750933.
Veltman, B., D. Harpaz, S. Melamed, Z. Tietel, L. Tsror, and E. Eltzov. 2022. “Whole-Cell Bacterial Biosensor for Volatile Detection From Pectobacterium-Infected Potatoes Enables Early Identification of Potato Tuber Soft Rot Disease.” Talanta247: 123545. https://doi.org/10.1016/j.talanta.2022.123545.
Vidal, J. E., H. P. Ludewick, R. M. Kunkel, D. Zähner, and K. P. Klugman. 2011. “The LuxS-Dependent Quorum-Sensing System Regulates Early Biofilm Formation by Streptococcus pneumoniae Strain D39.” Infection and Immunity79, no. 10: 4050–4060. https://doi.org/10.1128/IAI.05186-11.
Vieira, F. J. D., P. Nadal-Jimenez, L. Teixeira, and K. B. Xavier. 2020. “Erwinia carotovora Quorum Sensing System Regulates Host-Specific Virulence Factors and Development Delay in Drosophila melanogaster.” mBio11, no. 3: e01292-20. https://doi.org/10.1128/mBio.01292-20.
Vinodhini, V., and M. Kavitha. 2024. “Deciphering agr Quorum Sensing in Staphylococcus aureus: Insights and Therapeutic Prospects.” Molecular Biology Reports51, no. 1: 155. https://doi.org/10.1007/s11033-023-08930-3.
Voyich, J. M., C. Vuong, M. DeWald, et al. 2009. “The SaeR/S Gene Regulatory System Is Essential for Innate Immune Evasion by Staphylococcus aureus.” Journal of Infectious Diseases199, no. 11: 1698–1706. https://doi.org/10.1086/598967.
Wang, D., F. Cui, L. Xi, X. Tan, J. Li, and T. Li. 2023. “Preparation of a Multifunctional Non-Stick Tamarind Polysaccharide–Polyvinyl Alcohol Hydrogel Immobilized With a Quorum Quenching Enzyme for Maintaining Fish Freshness.” Carbohydrate Polymers302: 120382. https://doi.org/10.1016/j.carbpol.2022.120382.
Wang, L., C. Hu, and L. Shao. 2017. “The Antimicrobial Activity of Nanoparticles: Present Situation and Prospects for the Future.” International Journal of Nanomedicine12: 1227–1249. https://doi.org/10.2147/IJN.S121956.
Wang, Y., B. Liu, D. Grenier, and L. Yi. 2019. “Regulatory Mechanisms of the LuxS/AI-2 System and Bacterial Resistance.” Antimicrobial Agents and Chemotherapy63, no. 10: e01186-19. https://doi.org/10.1128/AAC.01186-19.
Waters, C. M., W. Lu, J. D. Rabinowitz, and B. L. Bassler. 2008. “Quorum Sensing Controls Biofilm Formation in Vibrio cholerae Through Modulation of Cyclic Di-GMP Levels and Repression of vpsT.” Journal of Bacteriology190, no. 7: 2527–2536. https://doi.org/10.1128/JB.01756-07.
Watson, W. T., T. D. Minogue, D. L. Val, S. B. von Bodman, and M. E. A. Churchill. 2002. “Structural Basis and Specificity of Acyl-Homoserine Lactone Signal Production in Bacterial Quorum Sensing.” Molecular Cell9, no. 3: 685–694. https://doi.org/10.1016/s1097-2765(02)00480-x.
Weiner, L. M., A. K. Webb, B. Limbago, et al. 2016. “Antimicrobial-Resistant Pathogens Associated With Healthcare-Associated Infections: Summary of Data Reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2011–2014.” Infection Control & Hospital Epidemiology37, no. 11: 1288–1301. https://doi.org/10.1017/ice.2016.174.
Whitehead, N. A., A. M. L. Barnard, H. Slater, N. J. L. Simpson, and G. P. C. Salmond. 2001. “Quorum-Sensing in Gram-Negative Bacteria.” FEMS Microbiology Reviews25, no. 4: 365–404. https://doi.org/10.1111/j.1574-6976.2001.tb00583.x.
Whiteley, M., S. P. Diggle, and E. P. Greenberg. 2018. “Erratum: Corrigendum: Progress in and Promise of Bacterial Quorum Sensing Research.” Nature555, no. 7694: 126. https://doi.org/10.1038/nature25977.
Wiechmann, A., V. Garcia, L. Elton, P. Williams, and S. Atkinson. 2023. “Reciprocal Regulation of NagC and Quorum Sensing Systems and Their Roles in hmsHFRS Expression and Biofilm Formation in Yersinia pseudotuberculosis.” Microbiology169, no. 10: 001397. https://doi.org/10.1099/mic.0.001397.
Winson, M. K., M. Camara, A. Latifi, et al. 1995. “Multiple N-Acyl-l-Homoserine Lactone Signal Molecules Regulate Production of Virulence Determinants and Secondary Metabolites in Pseudomonas aeruginosa.” Proceedings of the National Academy of Sciences92, no. 20: 9427–9431. https://doi.org/10.1073/pnas.92.20.9427.
Winzer, K., C. Falconer, N. C. Garber, S. P. Diggle, M. Camara, and P. Williams. 2000. “The Pseudomonas aeruginosa Lectins PA-IL and PA-IIL Are Controlled by Quorum Sensing and by RpoS.” Journal of Bacteriology182, no. 22: 6401–6411. https://doi.org/10.1128/JB.182.22.6401-6411.2000.
Wisniewski-Dyé, F., and J. A. Downie. 2002. “Quorum-Sensing in Rhizobium.” Antonie Van Leeuwenhoek81, no. 1–4: 397–407. https://doi.org/10.1023/a:1020501104051.
Woelber, J. P., A. Al-Ahmad, and K. W. Alt. 2022. “On the Pathogenicity of the Oral Biofilm: A Critical Review From a Biological, Evolutionary, and Nutritional Point of View.” Nutrients14, no. 10: 2174. https://doi.org/10.3390/nu14102174.
Wood, D. W., and L. S. Pierson, 3rd. 1996. “The phzI Gene of Pseudomonas aureofaciens 30–84 Is Responsible for the Production of a Diffusible Signal Required for Phenazine Antibiotic Production.” Gene168, no. 1: 49–53. https://doi.org/10.1016/0378-1119(95)00754-7.
Wood, D. W., F. Gong, M. M. Daykin, P. Williams, and L. S. Pierson, 3rd. 1997. “N-Acyl-Homoserine Lactone-Mediated Regulation of Phenazine Gene Expression by Pseudomonas aureofaciens 30–84 in the Wheat Rhizosphere.” Journal of Bacteriology179, no. 24: 7663–7670. https://doi.org/10.1128/jb.179.24.7663-7670.1997.
Wu, Y., C. W. Wang, D. Wang, and N. Wei. 2021. “A Whole-Cell Biosensor for Point-of-Care Detection of Waterborne Bacterial Pathogens.” ACS Synthetic Biology10, no. 2: 333–344. https://doi.org/10.1021/acssynbio.0c00491.
Wynendaele, E., N. Debunne, Y. Janssens, et al. 2022. “The Quorum Sensing Peptide Entf* Promotes Colorectal Cancer Metastasis in Mice: A New Factor in the Host–Microbiome Interaction.” BMC Biology20, no. 1: 151. https://doi.org/10.1186/s12915-022-01317-z.
Xu, L., H. Li, C. Vuong, et al. 2006. “Role of the luxS Quorum-Sensing System in Biofilm Formation and Virulence of Staphylococcus epidermidis.” Infection and Immunity74, no. 1: 488–496. https://doi.org/10.1128/IAI.74.1.488-496.2006.
Xu, X., S. Wang, C. Li, J. Li, F. Gao, and L. Zheng. 2023. “Quorum Sensing Bacteria in Microplastics Epiphytic Biofilms and Their Biological Characteristics Which Potentially Impact Marine Ecosystem.” Ecotoxicology and Environmental Safety264: 115444. https://doi.org/10.1016/j.ecoenv.2023.115444.
Xue, T., L. Zhao, and B. Sun. 2013. “LuxS/AI-2 System Is Involved in Antibiotic Susceptibility and Autolysis in Staphylococcus aureus NCTC 8325.” International Journal of Antimicrobial Agents41, no. 1: 85–89. https://doi.org/10.1016/j.ijantimicag.2012.08.016.
Yang, L., Y. Liu, C. Sternberg, and S. Molin. 2010. “Evaluation of Enoyl-Acyl Carrier Protein Reductase Inhibitors as Pseudomonas aeruginosa Quorum-Quenching Reagents.” Molecules15, no. 2: 780–792. https://doi.org/10.3390/molecules15020780.
Yang, L., K. B. Barken, M. E. Skindersoe, A. B. Christensen, M. Givskov, and T. Tolker-Nielsen. 2007. “Effects of Iron on DNA Release and Biofilm Development by Pseudomonas aeruginosa.” Microbiology153, no. Pt 5: 1318–1328. https://doi.org/10.1099/mic.0.2006/004911-0.
Yehuda, A., L. Slamti, R. Bochnik-Tamir, E. Malach, D. Lereclus, and Z. Hayouka. 2018. “Turning Off Bacillus cereus Quorum Sensing System With Peptidic Analogs.” Chemical Communications54, no. 70: 9777–9780. https://doi.org/10.1039/c8cc05496g.
Yu, D., L. Zhao, T. Xue, and B. Sun. 2012. “Staphylococcus aureus autoinducer-2 Quorum Sensing Decreases Biofilm Formation in an icaR-Dependent Manner.” BMC Microbiology12: 288. https://doi.org/10.1186/1471-2180-12-288.
Yu, X., B. Zhang, L. Zhang, and X. Wu. 2022. “The Regulatory Network Involving PcoR, RsaL, and MvaT Coordinates the Quorum-Sensing System in Pseudomonas fluorescens 2P24.” Applied and Environmental Microbiology88, no. 13: e0062522. https://doi.org/10.1128/aem.00625-22.
Zan, J., Y. Liu, C. Fuqua, and R. Hill. 2014. “Acyl-Homoserine Lactone Quorum Sensing in the Roseobacter Clade.” International Journal of Molecular Sciences15, no. 1: 654–669. https://doi.org/10.3390/ijms15010654.
Zettler, E. R., T. J. Mincer, and L. A. Amaral-Zettler. 2013. “Life in the “Plastisphere”: Microbial Communities on Plastic Marine Debris.” Environmental Science & Technology47, no. 13: 7137–7146. https://doi.org/10.1021/es401288x.
Zhang, H. B., L. H. Wang, and L. H. Zhang. 2002. “Genetic Control of Quorum-Sensing Signal Turnover in Agrobacterium tumefaciens.” Proceedings of the National Academy of Sciences99, no. 7: 4638–4643. https://doi.org/10.1073/pnas.022056699.
Zhang, M., C. A. Swofford, and N. S. Forbes. 2014. “Lipid A Controls the Robustness of Intratumoral Accumulation of Attenuated Salmonella in Mice.” International Journal of Cancer135, no. 3: 647–657. https://doi.org/10.1002/ijc.28700.
Zhang, W., H. Lu, W. Zhang, et al. 2024. “Inflammatory Microenvironment-Responsive Hydrogels Enclosed With Quorum Sensing Inhibitor for Treating Post-Traumatic Osteomyelitis.” Advanced Science11, no. 20: e2307969. https://doi.org/10.1002/advs.202307969.
Zhang, Y., A. Sass, H. Van Acker, et al. 2018. “Coumarin Reduces Virulence and Biofilm Formation in Pseudomonas aeruginosa by Affecting Quorum Sensing, Type III Secretion and C-di-GMP Levels.” Frontiers in Microbiology9: 1952. https://doi.org/10.3389/fmicb.2018.01952.
Zibafar, E., S. J. Hashemi, F. Zaini, H. Zeraati, S. Rezaie, and P. Kordbacheh. 2009. “Inhibitory Effect of Farnesol on Biofilm Formation by Candida tropicalis.” Daru-Journal of Pharmaceutical Sciences17, no. 1: 19–23.
Please check your email for instructions on resetting your password.
If you do not receive an email within 10 minutes, your email address may not be registered,
and you may need to create a new Wiley Online Library account.
Request Username
Can't sign in? Forgot your username?
Enter your email address below and we will send you your username
If the address matches an existing account you will receive an email with instructions to retrieve your username
The full text of this article hosted at iucr.org is unavailable due to technical difficulties.