

Allometry and spatial scales of foraging in mammalian herbivores
Abstract
Ecology Letters (2010) 13: 311–320
Herbivores forage in spatially complex habitats. Due to allometry and scale-dependent foraging, herbivores are hypothesized to perceive and respond to heterogeneity of resources at scales relative to their body sizes. This hypothesis has not been manipulatively tested for animals with only moderate differences in body size and similar food niches. We compared short-term spatial foraging behavior of two herbivores (sheep and cattle) with similar dietary niche but differing body size. Although intake rates scaled allometrically with body mass (mass0.75), spatial foraging strategies substantially differed, with cattle exhibiting a coarser-grained use of the ‘foodscape.’ Selectivity by cattle (and not sheep) for their preferred food was more restricted when patches were smaller (< 10 m2). We conclude that differences in spatial scales of selection offers a plausible mechanism by which species can coexist on shared resources that exhibit multiple scales of spatial heterogeneity.
Introduction
Spatial heterogeneity of food resources can result from aggregations of plants and nutrients, so that seemingly continuous pastures are actually highly variable with respect to nutrient availability (Wiens 1976; Kotliar & Wiens 1990; Searle et al. 2005). ‘Patchiness,’ occurs in pastures on scales ranging from centimeters to kilometers. The mechanisms by which herbivores perceive and respond to such patchiness remain uncertain, but it is clear that mammalian herbivores demonstrate high variability in spatial patterns of resource use (Bailey et al. 1996; Wilmshurst et al. 1999, 2000; Fryxell et al. 2005).
Ecologists have theorized that scaling relationships may lead to herbivore niche separation over evolutionary time among species utilizing shared, heterogeneous resources. These theories are based on physiologic and digestive consequences of body size, and apportion resources on a chemical axis (fiber fractions) based on body mass (e.g. Bell 1970; Jarman 1974). This is because allometric scaling laws predict constraints on metabolic demand, stride length, dental arcade width, and rumen volume, which could lead to different costs and limitations on nutrient intake, search, and movement. Ultimately, this could provide a mechanism by which larger animals may be restricted to taller, lower quality food options because, for example, larger species can utilize more fibrous foods, while smaller species are more capable of selecting less abundant food items of higher quality (Illius & Gordon 1987).
Resource exploitation at different spatial scales in fractal landscapes also predicts niche separation among species of different sizes, and offers an additional mechanism for coexistence of herbivores on shared resources (Ritchie 1998; Ritchie & Olff 1999). Such spatially driven niche separation models rely on the assumption that the main scales (or grain size) at which animals perceive and respond to heterogeneity in their environments are bounded by limitations related to body size. Remarkably, this hypothesized mechanism postulates a completely new niche axis, the scale at which resources are evaluated and used by herbivores (Cromsigt & Olff 2006). To our knowledge, this mechanism has never been tested in a controlled and manipulative experiment, especially on a range of scales and body sizes that are only moderately different, and with animals that have very similar ‘chemical’ dietary niches.
If smaller herbivores are bounded by allometric scaling relationships and indeed perceive their environments, or the ‘foodscape,’ at finer scales (Searle et al. 2007), then foraging selectivity – defined as the ability to exhibit selection of a preferred forage when given two or more options of contrasting quality – should be most pronounced at the optimal scale (grain size) for each species. Yet, the differences in the scales of perception that might affect an animal’s selectivity of one food option over another are challenging to study, and thus, remain poorly understood.
Our study focuses on the effects of forage spatial distribution on short-term (minutes) herbivore selectivity measured at spatial scales spanning two orders of magnitude from the bite (10−1 m) to local patches (10 m). We investigate the grazing choices of two herbivores, different in body size by one order of magnitude, but with similar dietary niche: cattle (Bos taurus) and sheep (Ovis aries). For grazing trials, we use constructed experimental arenas where the spatial distribution of constant quantities of two plant species is controlled. We test the hypotheses that (1) coarser-scale clustering enhances herbivore selectivity, when compared with selectivity in more finely interspersed environments, (2) smaller herbivores are more selective at all scales of observation, (3) selectivity of smaller herbivores is maximal at smaller grain sizes compared with that of larger herbivores and (4) given the same heterogeneous landscape, foraging patterns of smaller herbivores corresponds with a finer-grained view of the ‘foodscape.’ This work contributes to our understanding of how differing body size can influence patterns of foraging, and whether spatial distribution of foods constitutes a dimension for niche differentiation in spatially heterogeneous environments.
Materials and methods
Fieldwork was performed at the Campo Experimental J. F. Villarino, Facultad de Ciencias Agrarias, Universidad Nacional de Rosario, Argentina (33.01° S. Lat.; 60.53° Long) over two experimental periods: period 1 from 31 May to 22 June 2000 and period 2 from 28 August to 7 September 2000.
Animals
A total of four adult, cross-bred, non-lactating ewes (Ovis aries) weighing 45–50 kg and four adult, lactating, Holstein cows (Bos taurus) weighing 570–630 kg were used for measurements. The animals were trained to the sampling routine and diet during 4 months before the experiments. During experimental periods, but between observations, animals grazed at a mixed alfalfa–fescue pasture close to the experimental arenas. The same animals were used for preference and selectivity measurements. To provide a suitable social environment for sheep, the experimental individual was accompanied by two flock members that were muzzled and could not forage.
The lactation state generally produces a weak but repeatable increase in preference for high-quality forage by both cattle and sheep (Parsons et al. 1994; Farruggia et al. 2006; Rutter 2006). The fact that the cattle used in this study – and not the sheep – were in lactation likely increased the similarity of the chemical dietary niches of these two herbivores (by increasing the cows’ preference for alfalfa beyond what would have been expected of cows in the non-lactating state), thereby increasing the comparability of these two species based on other factors such as body size.
Plants
Alfalfa (Medicago sativa) and fescue (Festuca arundinacea) were seeded and grown in 6-L pots in an outdoor nursery in March 2000. A total of 2500 plants were grown, watered regularly, and fertilized monthly with 30 kg ha−1 ammonium phosphate. Pots were staked to the ground to form experimental arenas described below. Pots grazed in any trial were moved back to the nursery and replaced with fresh ones prior to the following trial. At the end of the first period, all plants were cut to even heights and returned to an outdoor nursery until the next period. Subsets of plants were collected and dried at 60 °C, and neutral-detergent fiber, acid detergent fiber and lignin were measured as described in Goering & Van Soest (1970).
Food preference
For initial preference trials, 10 pots of one plant species were arranged on the right side and 10 pots of the other species were on the left side of a corridor with a 0.5 m-wide lane between plant species. A session consisted of allowing each animal to take 20 bites. Each animal was tested once with alfalfa on the right side and fescue on the left, and then again with the orientation reversed in a complete randomized block design with two herbivore species × two sides and four replicates. Sessions were video-taped to determine the number of bites and foraging time per forage species. Preference for alfalfa was calculated as: (1) the number of bites of alfalfa divided by the total number bites taken, (2) the proportion of alfalfa mass in the diet consumed and (3) the percentage of the total time that animals spent eating alfalfa.
Selectivity
Definition of selectivity
Selectivity is defined as the relative proportion of a selected forage divided by the relative availability of that forage on the landscape (Senft 1989). For this study, we extended this concept by calculating selectivity for different hierarchical levels in the grazing process: the landscape (patches and paths), feeding stations (nodes encountered and pots grazed), bites, and intake (grams of dry matter, g DM). By doing so, we were able to elucidate the specific scales at which herbivores exhibited diet choices: by moving into and staying in preferred forage patches, by approaching nodes (at preferred feeding stations), by deciding how many bites to take in each pot, and by varying the size of bites.
Two selectivity indices were used, partial and cumulative selectivity (Table 1), similar to Wallis DeVries et al. (1999). Partial selectivity at a particular level (i.e. patches, paths, feeding stations, bites, or intake) was the percentage of preferred forage (alfalfa) selected at that level divided by the percentage available at that level (with availability determined by selection at higher levels; Table 1). Cumulative selectivity was calculated as the overall percentage of alfalfa selected at a particular level, compared with the availability of alfalfa on the total landscape. Under random selection, the expected percentage of each forage in the diet would be equivalent to that offered (cumulative selectivity) or encountered (partial selectivity) and both the partial and cumulative selectivities would be equal to 1.0. Values > 1.0 (< 1.0) indicated that the preferred forage was selected more (less) than expected by random encounter. Because the availability of alfalfa on the landscape was kept constant at c. 50% the maximum cumulative selectivity was c. 2.0 in our experiments. Otherwise, selectivity defined in this way can range between zero and infinity.
| Level | Parameter measured on Alfalfa | Parameter measured on fescue | Partial selectivity index | |
|---|---|---|---|---|
| 1 | Patch | # Patches entered, Pa | # Patches entered, Pf | S
Patch = 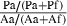 |
| 2 | Path length | Distance walked, Da | Distance walked, Df | S
Path = 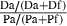 |
| 3 | Containers (pots; feeding stations) | # Containers with ≥ *1 bite, Ca | # Containers with ≥ *1 bite, Cf | S
Feeding station = 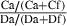 |
| 4 | Bites | # of bites, Ba | # of bites, Bf | S
Bite = 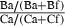 |
| 5 | Intake | g DM consumed, Ia | g DM consumed, If | S
Intake = 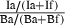 |
- Partial selectivity equations are shown. Cumulative selectivities at each level are calculated as the product of the partial selectivities at that level and at all higher (moving upward among the rows of the table) levels. For example, the cumulative selectivity at the feeding-station level equals SFeeding station × SPath × SPatch.
- g DM, grams of dry matter.
By studying partial selectivity, we were able to determine the forager’s ability to modify selectivity at smaller scales depending on the selectivity achieved at larger scales. Specifically, we tested whether sheep were better able than cattle to compensate in more homogeneous, finely interspersed environments (such as in the uniform treatment; Fig. 1a) by increasing the partial selectivity at smaller scales. Cumulative selectivity, on the other hand, was used to examine the overall result of simultaneous or consecutive choices compounded over multiple, hierarchical scales.

Experimental arenas: (a) uniform; (b) random-sized patches; (c) small patches; (d) large patches. Boundary lines show the edges of each arena, with circles marking nodes at which two 6 L pots were staked together. Solid circles = alfalfa; open circles = fescue; half-filled circles (in uniform arena only) = nodes where one pot each of alfalfa and fescue were both attached.
Selectivity experiments
Selectivity sessions took place in 40 × 40 m2 arenas containing plants arranged on a 3-m triangular lattice with 156 nodes, each having two pots marked by a coordinate label (for a total of 312 pots arranged in 12 columns × 13 rows; Fig. 1). Treatments were: (1) uniform – one pot of each species at each node, (2) random-sized patches – two pots with the same randomly chosen species at each node (the same random pattern was used throughout), (3) small patches – alternating monospecific clusters of six nodes with two pots per node and (4) large patches – alternate triangular monospecific clusters of 15 nodes. A factorial of four spatial patterns × two herbivore species × four animals in an incomplete randomized block (period) design was used. Combinations of factors were balanced between blocks (see Table S1).
Two selectivity sessions were carried out each day, starting in late morning or midday. Each animal was allowed to graze freely in the arena until 25% of the locations with potted plants (nodes) were encountered. All sessions were video-taped.
Animal paths were recorded on maps of the arenas, later verified using the video tapes, and then digitized to measure path lengths. A node was considered ‘encountered’ if the animal passed < 1.5 m from it, and ‘grazed’ if the animal ate any bites at the node. Observers also recorded the number of bites and residence time at each pot.
Biomass and dry matter intake calculations
Biomass removed per pot was estimated by weighing the fresh mass of plants and their pots at 1 g accuracy before and after each grazing bout. Six control pots of each species were kept under similar conditions near the experimental arenas and weighed every 30 min. These weights were later used to correct for fresh weight loss by evapo-transpiration. DM content of the consumed forage was determined by comparing DM of undefoliated vs. defoliated pots of each species.
Spatial analyzes and statistical tests
Hypotheses 1–3 were posed as linear combinations of estimated parameters and tested by bootstrapping 1999 samples. We used sampling with replacement of groups of observations to preserve patterns of correlations in residuals potentially induced by a random subject effect. Bootstrapped results were used to obtain mean absolute deviations around treatment means.
The maps of number of bites taken from each node in each session were analyzed by multi-scale pattern analysis (MSPA), an ordination method that partitions spatial variation into orthogonal scale components using Moran’s eigenvectors maps (MEM’s; Jombart et al. 2009). First, the set of 156 nodes is modeled by a network to obtain 155 orthogonal MEM’s (uj), each containing a set of weights for the nodes. The weights can be thought of as spatial ‘contrasts’ where, for example, the first eigenvector u1 compares bite density in the east vs. the west half of the grid, and u155 compares bite density in the white vs. black squares if the grid is visualized as a checkerboard. Regression of bite density against all 155 u’s produces a partition of its variation into 155 orthogonal spatial components, each representing a scale. The proportion of spatial variation of bites explained by each vector –R2 in mspa1 object (see appendix 1 of Jombart et al. 2009) – was summed within five scale classes (vectors 0–40, 41–80, 81–120, 121–140, and 141–155). The proportions explained by each class were then analyzed as repeated measures in a manova with herbivore and food distribution and their interaction as explanatory variables.
The ‘random-sized patch’ treatment (Fig. 1b) contained variability in the size of forage patches within the arena, and this was used to test whether spatial patterns of resource utilization varied with each herbivore’s scale of perception of the ‘foodscape.’ We graphed patches of alfalfa at different scales within the arena by convolution of a Gaussian kernel – of standard deviation sigma – with point masses at each alfalfa node, using the function ‘density.ppp’ in the R library ‘spatstat’ (R version 2.8.1, The R Foundation for Statistical Computing; http://www.r-project.org/). We varied sigma from 1 to 3 m (at 0.5 m increments) and used ordinal logistic regression to relate the resultant relative alfalfa ‘intensity’ at each node (low, medium, or high), computed at various scales (sigma), with the pattern of node utilization by the animals, measured as the number of sheep or cattle which grazed at that node in the selectivity trials.
Statistical tests to determine the effects of spatial pattern, herbivore species, experimental period, and their interactions were carried out using jmp version 8 (SAS Institute Inc., Cary, NC, USA) and spatial statistics were performed using the libraries ‘spatstat’ and ‘spdep’ in R version 2.8.1 (The R Foundation for Statistical Computing). When parametric tests were used, assumptions were checked, and transformations were used when necessary. When transformations were unsuccessful, nonparametric tests were used.
Results
Preference for alfalfa
There was a significant preference for alfalfa over fescue by both sheep and cattle (see Table S2 for details). Most of the animals chose to consume mixed diets (i.e. < 100% alfalfa). Sheep tended to show a stronger preference for alfalfa than cattle. The difference in preference between sheep and cattle was only marginally significant by Kruskal–Wallis rank sum tests (blocked by side offered) when measured by bites (P = 0.05) and DM intake (P = 0.05), although it was clearly significant by time (P = 0.01). Nevertheless, the trends for all three measures were in agreement. Bite-by-bite, in the preference trials, sheep were 1.2 (95% confidence interval, 1.1–1.4) times more likely to choose alfalfa than were cattle.
Overall foraging behavior
The total distance walked was limited because animals were allowed to graze until they had encountered c. 25% of the available nodes. Therefore, it is not surprising that the total distance walked was indistinguishable among sheep and cattle. However, most other measures of behavior during the grazing trials differed between these two herbivores (see Fig. S1 for details). Time spent consuming vegetation and total number of bites consumed were both significantly greater for sheep. Despite this, the average DM intake rate was greater for cattle, as expected because cattle took larger bites. Cattle were able to cover more ground in less time than sheep, probably due to their longer strides.
Intake rate corrected by metabolic weight did not differ significantly between sheep and cattle (see Fig. S1), meaning that DM intake rate scaled allometrically with body mass. None of the measures of grazing behavior – including intake/bite, bites/pot, feeding stations-grazed/m, m/s, total time spent consuming vegetation, total number of bites consumed, and total distance walked – differed significantly among the four experimental landscape patterns for sheep or cattle, with one exception: the average number of pots grazed per node encountered was lower in the uniform spatial arrangement for sheep only. This was because, in the uniform arena, sheep generally avoided fescue pots, meaning that they generally grazed at only one pot per node encountered. This was not so in the other treatments, where both pots in each node contained the same species.
Partial and cumulative selectivity
Considering all levels for the partial and cumulative indices (so that patch-, path-, feeding-station-, bite-, and intake-level selectivities were taken together as a multivariate response by manova), there was a significant effect of herbivore species (sheep vs. cow) on both the partial (P < 0.0001) and cumulative (P < 0.0001) selectivities. There was also a significant effect of landscape pattern (large patch, random-sized patch, small patch, or uniform) on both the partial (P = 0.0003) and cumulative (P = 0.0026) selectivity indices.
Landscape pattern and herbivore species had a significant interaction for the partial (P = 0.01) but not the cumulative (P = 0.09) selectivity indices. That is, changes in partial selectivity differed among the two species of herbivore as the spatial arrangement of forage changed. As the forages became more finely interspersed, sheep increased their partial selectivities at the feeding-station level (Fig. 2b). Thus, sheep achieved nearly as much cumulative selectivity in fine-grained as coarse-grained heterogeneous environments (see ‘cumulative intake’; Fig. 2d). For cattle, selectivity was maximal at the patch level and in the large patch treatment, and was restricted as forages became more finely interspersed, in the random-sized patch, small patch, and uniform environments (Fig. 2a). Cumulatively, cattle were unable to exhibit any selectivity in the small patch and uniform patterns (Fig. 2c). Thus, in contrast to sheep, which adjusted their behavior in finer-grained environments by increasing their selectivity at finer-scales, cattle were restricted and exhibited very little to no selectivity in those environments.

The effect of landscape configuration of forages on partial selectivity for a) cattle and b) sheep and cumulative selectivity for c) cattle and d) sheep within experimental arenas: large patches, random-sized patches, small patches, and uniform (as in Fig. 1). In panels a–b, the expected preferences are shown for both cattle and sheep, calculated from the results of independent preference trials (labeled gray arrows). Lines were smoothed to assist visualization, but data for each graph was collected only at the four discrete treatments. Bars are the mean absolute deviations calculated by nonparametric bootstrapping with 1999 bootstrap samples. Where bars are not visible, the deviations are so small as to fall completely behind the symbols.
There was a significant effect of period for both the partial (manova, P = 0.01) and the cumulative (manova, P = 0.006) selectivities. Most of this effect was due to differences at the patch level, with both herbivores exhibiting slightly higher patch-level partial selectivity, on average, during period 2 (Kruskal–Wallis rank sum test, P = 0.006), when herbage mass was taller. Total DM available for both alfalfa and fescue in each arena was much higher during period 2, with nearly twice the DM for both forages compared with period 1. However, as both forages increased similarly, the percentages available of alfalfa : fescue, on a DM basis, remained similar between periods: 39%:61% in period 1, 46%:54% in period 2. The average characteristics of available forage, other than total biomass availability, were similar among periods (see Table S3).
Including individual-animal preference as a covariate in the multivariate analysis for partial and cumulative selectivities neither change the outcomes reported above for the other predictors (herbivore, landscape pattern and period), nor did it reveal a significant overall relationship between individual preference and partial (P = 0.89) nor cumulative (P = 0.79) selectivity. This suggests that some other factors associated with foraging decisions, besides individual variation in preference, are needed to account for the observed differences in selectivity among individuals. Time of the day at the start of each foraging session was also tested as a covariate for both partial and cumulative selectivity, but it was not significant (P > 0.10).
Scales of resource utilization
The bootstrapping tests showed that partial selectivity of sheep was significantly greater than that of cows at the bite, feeding station, path, and patch scales (P < 0.01), but not at the intake levels. Mean sheep – cow differences, from intake to patch scale, were 0.12, 0.15, 0.67, 0.94, and 0.27. Sheep’s partial selectivity was maximal at the feeding station (48% of bootstrap samples) and path (52%) scales, whereas cow’s selectivity peaked at the feeding station, path and patch scales in 45, 25, and 19% of the 1999 bootstrap samples.
The MSPA showed that most of the spatial pattern in bites was at scales > 100 m2, reflecting the pattern of areas visited vs. areas never reached by the animals during each grazing session. The manova detected a significant interaction between food distribution and herbivore in the scales of bite placement, whereby sheep favored the smallest group of scales (0–10 m2) more than cattle (Fig. 3), particularly in the uniform food distribution.

Canonical scores for the first axis of a manova where proportions of spatial variation of bite density across scales were analyzed as a function of animal species, spatial arrangement of food, and their interaction. The canonical scores displayed on the vertical axis are directly proportional to small scales and inversely related to large scales.
In the random-sized patch treatment, patch sizes could vary with the spatial scale of perception. The relative density of alfalfa at each node, or the ‘intensity’ of alfalfa (low, medium, and high; Fig. 4), was recorded at each node for all five scales of perception (sigma = 1, 1.5, 2, 2.5, and 3 m). Ordinal logistic regression, with cow (or sheep) utilization as the dependent variable and alfalfa intensity at each of the five scales as the predictor variables, revealed that sheep utilization at each node was more strongly associated with the intensity of alfalfa at sigma = 1 m than with sigma =1.5, 2, 2.5, or 3 m. Conversely, cattle utilization was associated with the intensity of alfalfa defined at sigma =3 m but not 1, 1.5, 2, or 2.5 m (see Table S4 for likelihood ratio results). This suggests that cattle perceived and used relatively larger patches (even though of lower average quality) than sheep, despite the fact that all animals were presented with the same, random spatial arrangement of forage options (Fig. 4).

Spatial grazing patterns of (a) cattle and (b) sheep compared with ‘patches’ of alfalfa determined (independently of grazing observations) by convolution of the isotropic Gaussian kernel of standard deviation sigma with point masses at each of the nodes containing alfalfa (using function ‘density.ppp’ in the R library ‘spatstat’) – with sigma set to 3 m in (a), 1 m in (b). Circle sizes represent the intensity of node utilization (or the number of animals that grazed at each node during selectivity sessions): points indicate nodes at which no animals grazed; circles (from smallest to largest) indicate nodes at which 1, 2, 3, or all 4 animals grazed. Colors represent the spatial distribution of alfalfa ‘intensity’ within the random-sized patch arena, i.e. the relative density of alfalfa over space, defined at the hypothetical scale of perception for each herbivore (see text for details): lowest, medium, and highest intensity areas are represented as blue, green, or yellow, respectively – the legends show corresponding alfalfa ‘intensity’ values. In grayscale: blue = darkest gray; green = medium gray; yellow = lightest gray.
Discussion
This study provides empirical evidence based on a manipulative experiment that the scale of patchiness differentially influences forage selectivity among herbivores with differing body size and similar dietary niche. Results support our hypotheses by showing that: (1) coarser-scale clustering enhances selectivity of both sheep and cattle, (2) smaller herbivores (sheep) exhibit higher selectivity than larger ones (cattle) at all scales of observation, (3) the partial selectivity of sheep is maximal at smaller scales than cattle and (4) the spatial pattern of grazing by sheep corresponds to finer-scale patches than cattle.
Collectively, these results suggest that smaller-bodied herbivores (sheep) perceive or respond to spatial heterogeneity at finer scales compared with larger-bodied herbivores (cattle). However, herbivores clearly exhibit selectivity at multiple scales simultaneously, highlighting the hierarchical nature of foraging behavior. Our experiments employed only two species of herbivore, which limits generalization to other species. Nevertheless, the results indicate that spatial scaling laws and body size may indeed be important determinants of herbivore resource use in spatially heterogeneous environments. These findings have important ramifications for both management of large herbivores and foraging theory.
Scale as a niche dimension
The diversity of apparently coexisting large herbivore species has captivated the attention of scientists for many decades. Given the principle of competitive exclusion, scholars sought explanations for the coexistence of these ‘competitors’ (Bell 1970; Jarman & Sinclair 1979; Illius & Gordon 1992; Murray & Baird 2008). All explanations were based on one or more relationships among body size, metabolic requirements, digestive physiology, and mouth or dental arcade size. Since their inception, theories explained coexistence by resource partitioning on axes of chemical composition (plant species and parts) and height or abundance of herbage in implicitly homogeneous ‘foodscapes.’ Because they are adapted to different ranges of fiber content or density of forages (Demment & Van Soest 1985), potential competitors separate themselves over space or time (Gwynne & Bell 1968; McNaughton 1976; Cromsigt & Olff 2006).
Our results support the hypothesis that scale of foraging is an additional niche dimension over which large herbivores can partition their resources (Milne 1992; Ritchie 1998; Cromsigt & Olff 2006). Both cattle and sheep achieved the highest cumulative selectivity when plants were aggregated into the largest patches. Within the range of patch sizes tested (10−2 m2 to 102 m2), both herbivores perceived the largest patches of preferred forage and preferentially used them. However, as forage options became more finely interspersed, sheep readily compensated by increasing selectivity at finer scales, while cattle behaved as though these highly interspersed, small patches were part of a single large, homogeneous patch, and thus consumed each forage species at random in proportion to its availability (Fig. 2). The fact that sheep bite placement presented a significantly stronger small-scale component than cattle in the uniform food distribution further suggests that smaller herbivores exhibit a finer pattern of foraging, even when the food distribution presents no pattern at larger scales (Fig. 3).
Sheep exhibited much higher partial selectivity compared with cattle at the path and feeding-station levels, indicating that sheep have a more acute ability, or drive, to discriminate at these scales, regardless of the landscape distribution of forages. This suggests that sheep have a slightly different foraging strategy than cattle, focusing on higher quality options at a finer grain even at scales much greater than those where mouth size and morphology are relevant (Allden & Whittaker 1970; Shipley et al. 1994; Gregorini et al. 2007; Shipley 2007). Sheep were also able to achieve higher cumulative selectivity than cattle at all tested scales of patchiness. This tendency for smaller herbivores to more strongly focus on higher quality food is consistent with the early work of Jarman (1974) and Bell (1970), and is in agreement with theoretic predictions (Illius & Gordon 1992; Belovsky 1997) and the results of the preference trials in this study.
Overall, our results demonstrate that differential utilization of forage resources by herbivores of differing body size can result from scale-dependent limitations on foraging selection among resource patches, in addition to differences in bite-scale behaviors, such as bite size or bite depth, as has been previously proposed. For example, explanations of differential resource utilization among grazing ruminants have, in the past, relied on mechanisms relating to allometric constraints on bite size (Shipley et al. 1994) or bite depth (Allden & Whittaker 1970), or both (Illius 1989). The threshold patch size at which cattle no longer exhibited selectivity in our experiments coincided with patch areas of c. 10 m2. A roughly circular patch of similar area would have a radius equivalent to the body length of an average dairy cow (c. 2 m). This may explain, at least in part, why environments with patches of this or smaller area were treated as homogeneous by cattle, because perceived costs of frequent turning may have outweighed the perceived benefits for this large-bodied herbivore. This finding is in agreement with Wallis DeVries et al. (1999) who tested selectivity of cattle on tall-low-quality vs. short-high-quality swards and found that cattle were much more selective in experimental pastures made up of alternating 25 m2 patches than in 4 m2 patches.
Most models from foraging theory (reviewed by Stephens & Krebs 1986) are not explicitly spatial, and scaling issues have frequently been ignored (Wallis DeVries et al. 1999). Our results suggest, however, that all else being equal, the spatial arrangement of plants across scales can strongly affect the foraging behavior of both sheep and cattle. This is consistent with the findings of other studies which have demonstrated important effects of patch size and plant aggregation on diet selection by domesticated and wild ruminants (Clarke et al. 1995; WallisDeVries et al. 1999; Dumont et al. 2002), although rarely have the effects of body size and spatial scale been considered simultaneously in an experimental setting.
The observed differences in scales of selectivity among herbivores of different sizes could be a mechanism driving niche separation because patches that are not utilized by large herbivores, due to insufficient size or resource density, may contain several smaller patches with sufficient resource density to be selected by smaller herbivores. Fine-scale interspersion of high and low quality food options could exclude larger herbivores due to their inability, or reluctance, to pick out higher quality resources at finer scales (restricting the availability of forage for larger herbivores), as apparently occurred in the small patch and uniform arenas for cattle (but not sheep) in our experiments.
In summary, our study represents rare empirical evidence in support of a recently proposed mechanism for coexistence of large herbivores: resource partitioning by grazers in heterogeneous landscapes can result from differential spatial scales of perception and selection (Ritchie & Olff 1999; Cromsigt & Olff 2006). Our experiments only considered short-term (minutes) grazing behavior to minimize the potential for confounding factors. Cumulative and long-term effects of differential scales of selection remain unknown. The effects of spatial heterogeneity and allometric scaling on long-term grazing behavior need to be tested, as emergent patterns may occur over longer temporal scales. Ultimately, foraging decisions reflect trade-offs among competing goals as well as perceived costs based on sensory information integrated over multiple scales simultaneously (Illius & Gordon 1993; Laca & Ortega 1996). Our results suggest that differential spatial scales of selectivity among foragers of differing body size offers a plausible mechanism behind ecological coexistence of organisms utilizing shared heterogeneous resources, and this warrants further work elucidating how these spatial scaling factors interact with other determinants of foraging behavior.
Acknowledgements
The authors thank Maximo Alonso, Gregory Galin, and Evan Girvetz for assistance. This research was partially funded by USDA award ARS-6293-CG 9701033.

