

Uncovering hidden genetic variation in photosynthesis of field-grown maize under ozone pollution
Abstract
Ozone is the most damaging air pollutant to crops, currently reducing Midwest US maize production by up to 10%, yet there has been very little effort to adapt germplasm for ozone tolerance. Ozone enters plants through stomata, reacts to form reactive oxygen species in the apoplast and ultimately decreases photosynthetic C gain. In this study, 10 diverse inbred parents were crossed in a half-diallel design to create 45 F1 hybrids, which were tested for ozone response in the field using free air concentration enrichment (FACE). Ozone stress increased the heritability of photosynthetic traits and altered genetic correlations among traits. Hybrids from parents Hp301 and NC338 showed greater sensitivity to ozone stress, and disrupted relationships among photosynthetic traits. The physiological responses underlying sensitivity to ozone differed in hybrids from the two parents, suggesting multiple mechanisms of response to oxidative stress. FACE technology was essential to this evaluation because genetic variation in photosynthesis under elevated ozone was not predictable based on performance at ambient ozone. These findings suggest that selection under elevated ozone is needed to identify deleterious alleles in the world's largest commodity crop.
1 INTRODUCTION
Developing crops that can be more productive under stressful growing conditions is a high priority for agriculture today, and will be increasingly necessary if we are to avoid production losses to climate change (Challinor et al., 2014; Lesk, Rowhani, & Ramankutty, 2016; Lobell et al., 2014). Traditionally, field trials under more extreme environmental conditions than are typical for major crop growing regions have been used to test germplasm developed by breeding or biotechnology. However, some stressors—such as ozone (O3) pollution—are too heterogeneous and unpredictable in time or space to make this approach feasible (Ainsworth, Rogers, & Leakey, 2008). In addition, future climate change will result in growing environments with elevated [CO2] and temperature for which there is no present-day analogue (Battisti & Naylor, 2009; Leakey & Lau, 2012). Controlled environment growth facilities can provide valuable information on genetic variation in crop responses to stress treatments and the mechanisms underlying genetic variation (Brosché et al., 2010; Burton, Burkey, Carter, Orf, & Cregan, 2016; Frei, Tanaka, & Wissuwa, 2008; Ueda, Siddique, & Frei, 2015), but results of such controlled environment experimentation do not always translate into improved performance under production conditions in the field (Ainsworth, Beier, et al., 2008; Araus & Cairns, 2014; McKersie, Bowley, & Jones, 1999; Passioura, 2012). Free-air concentration enrichment (FACE) was developed to expose crops under field conditions to elevated concentrations of atmospheric pollutants over the entire growing season, with little or no perturbation to other aspects of the environment (Long, Ainsworth, Rogers, & Ort, 2004). But, most FACE experiments have tested a limited number of genotypes, at any given time (Betzelberger et al., 2010; Markelz, Strellner, & Leakey, 2011; Wang et al., 2014). Investigation of many genotypes and structured populations is needed to understand the heritability of traits in altered atmospheric environments and ultimately to identify genomic regions and genes associated with O3 tolerance.
Tropospheric O3 is a dynamic, short-lived air pollutant that is estimated to cause annual losses of ~10% to US maize yields with crop losses of $7.2 billion (McGrath et al., 2015). However, crop yield losses to O3 pollution are not widely recognized by farmers. And, breeding or biotechnology for tolerance to O3 stress has not been a major target for seed companies (Ainsworth, 2017). Ozone is formed as a secondary pollutant from nitrogen oxides (NOx) and volatile organic compounds, and recent analyses suggest that progress towards reducing NOx in the United States has slowed considerably, thus increasing the risks of O3 pollution (Jiang et al., 2018). Ozone diffuses through stomatal pores on leaf surfaces and reacts to form reactive oxygen species (ROS) in the apoplast. When ROS exceed the antioxidant-quenching capacity of the apoplast, they cause oxidative stress within cells that accelerates senescence and impairs photosynthesis, ultimately reducing plant productivity and crop yields (Ainsworth, Yendrek, Sitch, Collins, & Emberson, 2012; Kangasjärvi, Jaspers, & Kollist, 2005). Maize, like many other crop species, is sensitive to O3 damage, and shows accelerated loss of photosynthetic capacity with continued exposure to the air pollutant (Fiscus, Brooker, & Burkey, 2005; Yendrek, Erice, et al., 2017). Additionally, stomatal closure can be negatively impacted by O3 stress, leading to excessive water loss under drought stress (Wang et al., 2014; Wilkinson & Davies, 2010). Maintenance of high photosynthetic CO2 assimilation without excess stomatal conductance is an important phenotype for increasing O3 tolerance (Ainsworth, 2017; Emberson et al., 2018). Furthermore, enhancing photosynthetic CO2 assimilation and water use efficiency (ratio of photosynthetic CO2 assimilation to water loss by transpiration) are widely recognized to be key targets for crop improvement at a time when potential for further gains in harvest index and planting density may be limited (Leakey et al., 2019; Long, Marshall-Colon, & Zhu, 2015; von Caemmerer & Furbank, 2016). Despite the importance of maize for food, fuel and animal feed, little is known about the extent or mechanisms of genetic variation in the sensitivity of maize to O3 by comparison to other crops such as soybean, wheat and rice (Betzelberger et al., 2012; Burton et al., 2016; Frei et al., 2008; Wang et al., 2014). This represents an important unexplored opportunity because maize is a highly tractable, model system for study of crop genetics (Buckler et al., 2009; Riedelsheimer et al., 2012; Schnable et al., 2009).
For physiological performance to be a target for improvement in breeding programmes, there must be underlying additive genetic variation in the traits of interest. The likely success of selection is reflected in the narrow sense heritability, that is, the proportion of phenotypic variation resulting from additive genetic variance (Falconer & Mackay, 1996; Flood, Harbinson, & Aarts, 2011). Previous studies of maize have estimated relatively high heritability for traits related to photosynthetic capacity (Cai et al., 2012; Crosbie, Mock, & Pearce, 1977; Lu et al., 2011; Pelleschi et al., 2006; Prado et al., 2017; Wang et al., 2013; Ziyomo & Bernardo, 2013) and indicated that variance in photosynthetic traits is mostly additive (Crosbie et al., 1977). But, the heritability of photosynthetic traits in crops is reportedly lower under stress conditions (Edwards, Ewers, McClung, Lou, & Weinig, 2012; Pelleschi et al., 2006). Prior studies on leaf-level responses to O3 in fescue, potato and sweetcorn found that additive effects (GCA), not dominance effects (SCA), were significant and involved in O3 tolerance (De Vos, Hill, Pell, & Cole, 1982; Johnston, Haaland, & Dickens, 1983; Schraudner, Langebartels, & Sandermann, 1997). These studies fumigated crops with very high concentrations of O3 for hours to days, which elicited acute stress responses that are known to be fundamentally distinct from responses to season-long, moderate O3 concentrations that drive yield loss in farmer's fields (Ainsworth et al., 2012; Chen, Frank, & Long, 2009; Schraudner et al., 1997). Uncertainty regarding the extent to which photosynthetic traits associated with O3 tolerance are heritable is compounded by the need to know if there are strong genetic correlations across environments (Falconer, 1952). In other words, if there is a substantial genotype × environment interaction acting on photosynthetic traits, then selection for crop genotypes than can tolerate elevated O3 pollution would not be successful under standard growing conditions. Alternatively, the absence of genotype × environment interaction would suggest that past selection for highly productive genotypes would likely have incidentally selected for tolerance to O3 pollution as well. In addition, genetic correlations among traits are useful in assessing how many independent traits need to be evaluated for a successful selection index to be developed.
Proof-of-concept is needed to demonstrate the use of FACE experimentation to estimate the heritability of photosynthetic traits and the degree to which elevated O3 affects heritability in a farm-field setting (Frei, 2015). Therefore, in this study, we used a half-diallel mating design to test for (a) the effects of elevated O3 on photosynthetic traits in maize; (b) the heritability and genetic correlations among photosynthetic traits in maize; and (c) the identification of particularly susceptible parental lines.
2 MATERIALS AND METHODS
2.1 Field site and experimental treatments
In 2016 and 2017, a maize half-diallel panel of 45 crosses among 10 maize inbred lines (Figure 1) was planted at the FACE field site located on the experimental farms of the University of Illinois at Urbana-Champaign (40°02′N, 88°14′W; www.igb.illinois.edu/soyface/). Seeds were planted on May 24, 2016 and May 17, 2017. Experimental rows were 3.35 m long with a planting density of 8 plants/m and row spacing of 0.76 m. Maize was grown in rotation with soybean, fertilized with N (200 kg/ha) and irrigated as needed using drip irrigation. Maize was exposed to ambient or elevated O3 in 20 m dia. octagonal rings (n = 4 paired ambient and elevated O3 rings). Each ring was divided into five sub-blocks of 10 rows each (Figure 1). The 45 genotypes were randomized into five entry sets of nine genotypes and entry-sets were randomly assigned to different sub-blocks in each of the four pairs of rings (one ambient, one elevated). Within each sub-block, B73 × Mo17 was planted as a check genotype.

The O3 treatment was applied at a target set point of 100 nl/L from 10:00 to 18:00 throughout the growing season when it was not raining, when leaves were dry and when wind speed was greater than 0.5 m/s (Yendrek, Erice, et al., 2017; Yendrek, Tomaz, et al., 2017). Based on 1 min average O3 concentration collected in each ring throughout the season, the fumigation was within 10% of the 100 nl/L target for 62% of the time and within 20% of the target concentration for 87% of the time in 2016. In 2017, fumigation was within 10% of the target for 59% of the time and within 20% of the target for 84% of the time. Other meteorological conditions were measured with an on-site weather station (Figure S1).
2.2 Measurements of photosynthetic traits
On July 5–9, 2016 and June 28–July 1, 2017 leaf gas exchange was measured. In 2016, the eighth leaf was measured (approximately the third or fourth fully expanded leaf), while in 2017 the third fully collared leaf was measured. Prior to dawn, leaves from two plants per genotype from one ambient and one elevated O3 plot were labelled and cut, providing ~208 leaves for measurements per day. Leaves were immediately placed in a bucket with the cut surface submerged in water, transported to a laboratory, recut under water and placed in 50 ml tubes filled with water. Leaves were stored in dim light (<100 µmol m−2 s−1). Approximately 1 hr before the measurement of leaf gas exchange, leaves were placed in a growth chamber with the following conditions: 1,800 photosynthetic photon flux density (PPFD, µmol m−2 s−1) at the leaf surface, 18°C air temperature (25°C leaf temperature) and relative humidity of ~90%. After approximately 1 hr of acclimation, leaf gas exchange was measured with a set of portable gas exchange systems (LI-6400; LI-COR Biosciences, Lincoln, NE, USA) with 2 × 3 red/blue LED cuvettes (LI-6400-02B; LI-COR Biosciences). Light-saturated gas exchange was measured at 1,800 PPFD (µmol m−2 s−1), 25°C (leaf temperature) and 60% relative humidity.
Gas exchange systems were run simultaneously, and leaf mean flux of net CO2 assimilation (A), stomatal conductance (gs) and the ratio of the leaf intercellular concentration of CO2 to atmospheric CO2 (ci:ca) were measured or calculated every 4 s for 4 min (Figure S2). Analysis was done on the average values of A, gs and ci:ca over the last minute of measurement. Instantaneous water use efficiency (iWUE = A/gs) was calculated from A and gs. Following gas exchange measurements, leaf reflectance was measured from the adaxial surface of the leaf using a full-range spectroradiometer (ASD FieldSpec 4 Standard Res, Analytical Spectral Devices) following a standard protocol (Yendrek, Erice, et al., 2017; Yendrek, Tomaz, et al., 2017). Six reflectance measurements were collected and a splice correction was applied to the spectra to ensure continuous data across detectors (Serbin, Singh, McNeil, Kingdon, & Townsend, 2014). Those six spectra from each leaf were then averaged and data were interpolated to provide 1 nm bandwidths. Further quality control was applied using the FieldSpectra package in R (Serbein et al., 2014). Partial least squares regression models were then applied to the spectra to estimate the CO2-saturated rate of A (Vmaxm) and leaf chlorophyll content (Chlm; Yendrek, Erice, et al., 2017; Yendrek, Tomaz, et al., 2017).
2.3 Data quality control
Measurements of gas exchange over time were examined for data quality and conformation to basic assumptions. If mean gs or ci for the last minute of the measurement period were negative, then data were not used (Figure S2). Individual leaf measurements for the same genotype and treatment were evaluated for outliers and influential observations. If an individual measurement strongly influenced the stability of the data, the observation was not used for further statistical analysis.
2.4 Statistical analysis

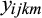 is the observed trait value for the ith treatment (ambient or elevated O3) in the kth sub-block of the jth ring-pair, belonging to the cross from the mth mother where µi is the mean for the ith treatment.
is the observed trait value for the ith treatment (ambient or elevated O3) in the kth sub-block of the jth ring-pair, belonging to the cross from the mth mother where µi is the mean for the ith treatment. 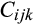 is the trait value of the B73 × Mo17 check plot in the ith treatment, jth ring-pair and kth sub-block.
is the trait value of the B73 × Mo17 check plot in the ith treatment, jth ring-pair and kth sub-block.  is the random GCA effect (parameterized as a set of indicator variables for the maternal parent). The variance component for GCA was estimated using a Toeplitz(1) variance structure (Wayne et al., 2007).
is the random GCA effect (parameterized as a set of indicator variables for the maternal parent). The variance component for GCA was estimated using a Toeplitz(1) variance structure (Wayne et al., 2007).
 estimated variance of GCA effect and
estimated variance of GCA effect and 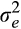 is residual variance. The SE of heritability estimates was obtained with the Delta method (Wayne et al., 2007).
is residual variance. The SE of heritability estimates was obtained with the Delta method (Wayne et al., 2007).


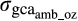 is the genetic covariance between environments for a given trait and
is the genetic covariance between environments for a given trait and 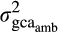 and
and 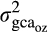 are the additive genetic variances for the same trait in ambient and elevated O3 respectively.
are the additive genetic variances for the same trait in ambient and elevated O3 respectively.


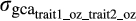 is the additive genetic covariance between traits for the same environment, and
is the additive genetic covariance between traits for the same environment, and 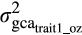 and
and  are the additive genetic variances for trait1 and trait2.
are the additive genetic variances for trait1 and trait2.
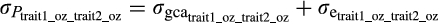 . All calculations were performed using code adapted from Holland (2006).
. All calculations were performed using code adapted from Holland (2006).Visual examination of the results led to the hypothesis that NC338 and HP301 were responsible for the shift in genetic correlations. Thus, the correlation analyses described above were repeated for the set of F1 hybrids that did not include hybrids NC338 or Hp301 as either parent.
3 RESULTS
3.1 Gas exchange response to elevated O3
Photosynthetic traits of 45 F1 hybrids comprising a half-diallel population were measured in ambient and elevated O3 in 2016 and 2017. Measurements were taken on mature leaves approximately midway through vegetative development of the crop in both years. Rainfall was lower in 2017, leading to greater drought stress than in 2016 (Figure S2), and lower A and gs (Figure 2; Figure S3). In both years, averaging across hybrids, elevated O3 reduced A, by ~14% (Figure 2), and gs by 6% in 2016 and by 14% in 2017 (Figure S3).

3.2 Narrow sense heritability increases with O3 environment
Analysis of the half-diallel maize population grown showed that additive genetic variance explained the variation in photosynthetic and reflectance traits (Table S1). There was little evidence for dominance variance (Table S1; comparison of models 2 and 3; Table S2). In addition, there was strong evidence for interactions between genetic variation and environment (G × E; Table S1; comparison of models 2 and 3), indicating that the genetic architecture of most traits was different in the two environments. Therefore, heritability was calculated separately in ambient and elevated O3. Narrow sense heritability (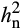 ) estimates of photosynthetic traits differed between ambient and elevated O3 in 2016 and 2017 (Table 1). Interestingly,
) estimates of photosynthetic traits differed between ambient and elevated O3 in 2016 and 2017 (Table 1). Interestingly, 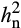 in elevated O3 was greater than in ambient O3 for many photosynthetic traits (Table 1), indicating the presence of additional additive genetic variation in elevated O3. Estimated
in elevated O3 was greater than in ambient O3 for many photosynthetic traits (Table 1), indicating the presence of additional additive genetic variation in elevated O3. Estimated 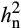 of A was greater than
of A was greater than 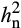 of other photosynthetic traits (Table 1), and was of similar magnitude to previous estimates in maize (Crosbie et al., 1977; Pelleschi et al., 2006).
of other photosynthetic traits (Table 1), and was of similar magnitude to previous estimates in maize (Crosbie et al., 1977; Pelleschi et al., 2006).
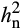 ) for photosynthetic traits measured in 2016 and 2017 in ambient and elevated [O3]
) for photosynthetic traits measured in 2016 and 2017 in ambient and elevated [O3]| 2016 | 2017 | |||
|---|---|---|---|---|
| Ambient O3 | Elevated O3 | Ambient O3 | Elevated O3 | |
| A | 0.222 | 0.678 | 0.315 | 0.499 |
| g s | 0.178 | 0.380 | 0.278 | 0.202 |
| iWUE | 0.127 | 0.182 | 0.212 | 0.203 |
| ci:ca | 0.103 | 0.182 | 0.197 | 0.237 |
| V maxm | 0.125 | 0.236 | 0.197 | 0.258 |
| Chlm | 0.424 | 0.283 | 0.171 | 0.276 |
3.3 Elevated O3 alters genetic and phenotypic correlations among photosynthetic traits
Genetic correlations (Rg) are important to define the shared genetic components between traits and phenotypic correlations (Rp) measure the consistency of performance between traits. Both Rg and Rp between A and gs were very strong regardless of the environment or year (Figure 3a,b, red symbols). This is unsurprising because gs is a principle determinant of CO2 entry into leaves, and A measures CO2 fixation. Rg between gs and iWUE were strongly negative in both 2016 and 2017, indicating that selection for high gs in maize would result in low iWUE (Figure 3, grey symbols). There was a positive Rg between traits estimated from leaf reflectance spectra, Chlm and Vmaxm (Figure 3a, olive symbols) in both ambient and elevated O3, but Rg between A and Vmaxm or Chlm was not strong or consistent (Figure 3a, pink symbols). It was notable that Rg between A and ci:ca (Figure 3a, green symbols) and gs and ci:ca (Figure 3a, cyan symbols) differed in ambient and elevated O3. Rg between gs and ci:ca was strong and positive in ambient O3, but not in elevated O3 (Figure 3a, cyan symbols), and Rg between A and ci:ca was positive in ambient O3 and negative in elevated O3 (Figure 3a, green symbols). These same general trends were also observed for Rp (Figure 3b).

3.4 Identification of O3-sensitive maize lines
The correlation between gs and ci:ca was further explored based on the differences in Rg between traits in ambient and elevated O3. Across the diverse F1 hybrids, there was a strong, linear relationship between gs and ci:ca consistent among all lines in ambient O3 (Figure 4, circles). However, growth at elevated O3 altered the relationship between gs and ci:ca in F1 crosses with parents NC338 (Figure 4, orange triangles) and Hp301 (Figure 4, blue triangles) in both 2016 and 2017. Notably, alleles from Hp301 and NC338 appear to confer sensitivity to elevated O3 in different ways. Lower A at elevated O3 in hybrids with Hp301 as a parent was linked to lower gs at elevated O3 without a change in ci:ca (Figure 4). Meanwhile, lower A at elevated O3 in hybrid with NC338 as a parent was linked to greater ci:ca without a change in gs (Figure 4). F1 hybrids with these two parents also showed the greatest per cent decrease in A and gs at elevated O3 in 2016 and 2017 (Figure 5a,b). When Rg and Rp were estimated without hybrids containing NC338 or Hp301 (Figure 3c,d), then estimates were similar in both ambient and elevated O3. This further indicates that F1 crosses with Hp301 and NC338 are sensitive to O3 and drive the differences in Rg and Rp observed between A and ci:ca and gs and ci:ca.


4 DISCUSSION
This 2-year study enabled investigation of genetic variation in photosynthetic traits as well as the consistency of the response of photosynthetic traits to elevated O3 across growing seasons. Photosynthesis has been suggested as one of the most important processes to improve in order to combat stagnating crop yields and improve future food security (Long et al., 2015). Transgenic approaches to improve C4 photosynthesis have been suggested (von Caemmerer & Furbank, 2016), but exploiting standing genetic variation in photosynthetic traits is also a path to crop improvement that does not involve transgenic technology (Cañas et al., 2017). Here, we showed that maize hybrids have significant additive genetic variation in A in both ambient and elevated O3 (Figure 2), and the per cent change in A at elevated O3 was consistent in both years (Figure 5a). By using a half-diallel mating design comprised of diverse maize lines, we estimated genetic (Rg) and phenotypic (Rp) correlations in both environments and demonstrated that growth at elevated O3 alters the Rg and Rp between photosynthetic traits. Growth at elevated O3 changes the relationship between gs and ci:ca, primarily driven by genotypes with Hp301 and NC338. Furthermore, the mechanisms of sensitivity to elevated O3 appear to differ between genotypes Hp301 and NC338. Overall, we have demonstrated the utility of FACE experiments for screening structured populations to estimate heritability of phenotypic traits and how those change with O3 pollution. This capacity and information is critical for any future efforts to breed for O3 tolerance.
Previous studies of O3-induced leaf damage in fescue, tobacco, maize, potato and Plantago major also showed little evidence of dominance variance (Aycock, 1972; Cameron, 1975; Campbell, Devine, & Howelp, 1977; De Vos et al., 1982; Huang, Aycock, & Mulchi, 1975; Johnston et al., 1983; Whitfield, Davison, & Ashenden, 1997), which is consistent with our findings. Lack of dominance variance indicates that the narrow sense heritability (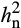 ) of net carbon assimilation (A) in elevated O3 is mainly additive and can be attributed to additive genetic factors inherited from the parents. The observed increase in
) of net carbon assimilation (A) in elevated O3 is mainly additive and can be attributed to additive genetic factors inherited from the parents. The observed increase in 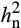 of A under O3 stress is notable. A prior study investigating O3 sensitivity in Black Cherry (Prunus serotine Ehrh.) also found that heritability of the foliar injury increased with exposure to greater treatment concentrations of elevated O3 (Lee, Steiner, Zhang, & Skelly, 2002). In contrast, studies that investigated other abiotic stresses such as drought stress often reported that
of A under O3 stress is notable. A prior study investigating O3 sensitivity in Black Cherry (Prunus serotine Ehrh.) also found that heritability of the foliar injury increased with exposure to greater treatment concentrations of elevated O3 (Lee, Steiner, Zhang, & Skelly, 2002). In contrast, studies that investigated other abiotic stresses such as drought stress often reported that 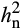 was lower under stress conditions. For example, drought stress decreased
was lower under stress conditions. For example, drought stress decreased 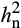 of A from 0.61 to 0.33 in Brassica rapa (Edwards et al., 2012) and from 0.45 to 0.37 in maize (Pelleschi et al., 2006). Greater
of A from 0.61 to 0.33 in Brassica rapa (Edwards et al., 2012) and from 0.45 to 0.37 in maize (Pelleschi et al., 2006). Greater 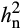 in elevated O3 as revealed in this study suggests that there is potential for improvement. Furthermore, selection for A under standard growing conditions of the Midwest in the United States would fail to select against deleterious alleles that confer sensitivity to elevated O3 (Gibson & Dworkin, 2004).
in elevated O3 as revealed in this study suggests that there is potential for improvement. Furthermore, selection for A under standard growing conditions of the Midwest in the United States would fail to select against deleterious alleles that confer sensitivity to elevated O3 (Gibson & Dworkin, 2004).
Genetic correlations (Rg) among traits imply that the same genes are acting on multiple traits. If the correlation is high enough, it is possible that selection can be performed on one trait, with the second trait also improving due to shared loci between the traits. We tested Rg among photosynthetic traits measured with gas exchange and traits estimated from leaf hyperspectral reflectance (Yendrek, Erice, et al., 2017; Yendrek, Tomaz, et al., 2017), which has been suggested as a promising high-throughput approach for phenotyping (Araus & Cairns, 2014; Furbank & Tester, 2011). Although Rg between Vmaxm and Chlm estimated from hyperspectral reflectance was strong and positive, Rg between gas exchange traits and remotely sensed traits was not strong in our experiments (Figure 3a). This could be because chlorophyll content and maximum photosynthetic capacity were not the primary limitations to gas exchange at the times of our measurement. In 2017, reflectance was measured in the field on intact plants, which may also lead to lower correlations. We did observe strong, positive correlations between A and gs, which has been reported previously for other species under variable environmental conditions (Manzaneda, Rey, Anderson, Raskin, & Mitchell, 2016; Pelleschi et al., 2006). We also identified a fundamental shift in Rg between gs and ci:ca in ambient and elevated O3 (Figure 3a). ci:ca provides information on the balance between resistance for CO2 diffusion into the leaf and the biochemical capacity for CO2 fixation in the mesophyll. Altered Rg in elevated O3 suggests the involvement of additional genetic factors in controlling the phenotype under O3 stress.
Further analysis of the relationship between gs and ci:ca identified that hybrids with NC338 and Hp301 were more sensitive to O3 stress (Figure 4). When Rg and Rp were calculated without hybrids containing Hp301 and NC338, the correlations become similar in ambient and elevated O3 (Figure 3c,d), suggesting that alleles from Hp301 and NC338 were responsible for the change in genetic architecture of photosynthetic traits in elevated O3. These alleles from Hp301 and NC338 demonstrate different mechanisms of sensitivity to elevated O3, with Hp301 crosses linked to lower gs at elevated O3 with no change in ci:ca and NC338 hybrids showing greater ci:ca without a change in gs (Figure 5). This implies that stomatal function was disrupted in Hp301 lines, which is consistent with the prior evidence that O3 stress alters stomatal signalling and closure (Wilkinson & Davies, 2010). Meanwhile, our findings suggest that alterations in mesophyll conductance (gm) or perhaps decreases in photosynthetic capacity drove sensitivity to elevated O3 in hybrids with NC338 as a parent. This is consistent with greater average reductions in Vmaxm and Chlm in hybrids containing NC338 as a parent (Figure 5). A study of Siebold's beech (Fagus crenata) seedlings found a significant increase in ci and a substantial decrease in gm with increasing O3 stress, suggesting that lower gm hindered CO2 transfer to the site of carbon fixation (Watanabe et al., 2018). Similarly, a study on snap peas found the genotype sensitive to elevated O3 had a significant decrease in gm (Flowers, Fiscus, Burkey, Booker, & Dubios, 2007). Photosynthetic processes may be sensitive to O3 (Fiscus et al., 2005) and reductions in PEPc activity and Rubisco activity were reported at elevated O3 in young maize leaves (Leitao, Bethenod, & Biolley, 2007). Such reductions in either gm or photosynthetic enzyme activity if decoupled from gs could increase ca, as observed in the sensitive NC338 hybrids here.
Global mean O3 pollution increased by 8.9% from 1990 to 2013 (Brauer et al., 2016) causing a significant, negative impact on crop production in North America and Europe worldwide (Mills et al., 2018). Identifying genetic variation in photosynthetic traits and tolerant genotypes to elevated O3 is a crucial step towards breeding for O3 tolerance (Ainsworth, 2017). For breeding to be straightforward, genetic variation should be directly available to selection, and the genetic contribution to phenotypic variation should be additive. This first investigation of a half-diallel population of maize exposed to elevated O3 in the field showed not only that additive genetic variation is a large component of phenotypic variation in photosynthetic traits, but also that O3 stress increased the heritability of photosynthetic traits, indicating additional loci responding to O3 stress. In addition, the genetic correlation among photosynthetic traits changed between environments. The F1 hybrids from parents Hp301 and NC338 were particularly sensitive to O3 and showed disrupted relationships between A or gs and ci:ca. The physiological mechanism underlying this sensitivity was distinct between the two parent lines, suggesting that multiple sources of sensitivity to oxidative stress from air pollution might need to be selected against to optimize maize production. This implies that past selection of maize under ambient O3 did not select for alleles that confer tolerance to elevated O3 pollution. Further selection under elevated O3 concentrations might purge deleterious alleles in the world's most important commodity crop. More broadly, this work demonstrates the capability of FACE technology to be used for field experimentation on diverse populations of major commodity crops to address needs for adaptation to pollution and climatic change that cannot be easily accomplished by other existing approaches.
ACKNOWLEDGEMENTS
This work was supported by a grant from the NSF Plant Genome Research Program (PGR-1238030). We thank Brad Dalsing, Chad Lance, Aidan McMahon and Jesse McGrath for operating fumigation at SoyFACE. We thank Chris Moller, Ben Thompson, John Ferguson, Charlie Burroughs, Lorena Rios-Acosta and Jessica Wedow for help with sampling, photosynthetic and reflectance measurements. We thank Jim Berry, Aya Bridgeland, Cecilia Pigozzi, Savannah Pflugmacher, Noah Mitchell and Emma Diers for their help in maintaining the research plots. We thank Linda Young for assistance with the spatial design of the experiment. Any opinions, findings and conclusions or recommendations expressed in this publication are those of the author(s) and do not necessarily reflect the views of the U.S. Department of Agriculture. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. USDA is an equal opportunity provider and employer.

