

Positive effect of spring advance on the diet quality of an alpine herbivore
Abstract
Changes in vegetation phenology related to global warming are having alarming effects on the life history traits of many herbivore species. Such changes are particularly critical in alpine ecosystems, where strong climate limitations on plant growth make seasonal synchronization imperative for the growth, reproduction and survival of herbivores. However, despite the pivotal role of resource-use strategies on the performances of such species, few studies have explicitly assessed the mechanistic impact of climate change on their diets. We aimed to fill this gap by studying the effect of spring onset on the dietary composition and quality of a medium-size alpine herbivore while considering density-dependent processes and age- and sex-specific differences in foraging behavior. Using an exceptional, long-term (24 years) direct individual-based dietary monitoring of a Pyrenean chamois population (Rupicapra pyrenaica pyrenaica), we showed that ongoing earlier onsets of spring are leading to an earlier access to high-quality forage and therefore a higher diet quality at a fixed date, without apparent changes in diet composition. We also showed that at high densities, intraspecific competition reduced diet quality by driving animals to feed more on woody plants and less on nutritious forbs and graminoids. By assessing the mechanistic effects of global warming on the dietary patterns of species at the center of trophic networks, this study is an essential step for predictive models aiming at understanding the ongoing ecosystem consequences of the global climatic crisis.
INTRODUCTION
Ongoing anthropogenic climate change is having dramatic effects on ecological systems and processes on all continents and across oceans (IPCC 2014), resulting in changes in the phenology, distribution and behavior of organisms (Parmesan 2006; Cleland et al. 2007). Yet, the direction and magnitude of these responses can differ and many underlying mechanisms are still unclear (Moritz & Agudo 2013; Stopher et al. 2014). It is therefore urgent to understand the mechanisms by which climate change is affecting the life history traits of organisms if we want to monitor, predict and mitigate future biodiversity changes (Sala et al. 2000).
Climate triggers numerous periodic events in the life of organisms (Mysterud et al. 2003) and the global increase in temperature is greatly affecting the seasonality of those events (Cleland et al. 2007). In temperate ecosystems, researchers are observing advancements of spring events such as the flowering of plants and the unfolding of leaves (Menzel 2000; Franks et al. 2007) and even the time of bird reproduction and migration (Charmantier & Gienapp 2014; Tomotani et al. 2018). However, the scarcity of long-term monitoring data and the difficulty to test multiple confounding factors hampers the assessment of these phenological shifts caused by climate change (Kharouba & Wolkovich 2020).
Alpine species are robust indicators of the ongoing changes that can be used to forecast their effects on the life-history traits of many organisms (Inouye & Wielgolaski 2013; Mason et al. 2014). Indeed, alpine ecosystems are among areas where climate warming is particularly intense, making alpine species as sentinels that may allow gaining critical insights on forthcoming effects expected elsewhere (Beniston 2005; EEA 2012). In addition, because mountain species face strong limiting environmental conditions for which they have developed unique adaptations, they are expected to be particularly vulnerable to climate change (IPCC 2014; White et al. 2018). For instance, the expected upward shift of trees due to climate warming constitutes a major threat to range-restricted high-altitude species, by shrinking the area of their suitable habitat (Dirnböck et al. 2011). By imposing intense seasonal changes in resource availability and quality (Post & Stenseth 1999; Inouye 2020), the strong climate limitation of plants growth in alpine ecosystems is also expected to have indirect effects on primary consumers, whose central place in trophic networks may broadcast the cascading ecological consequences of climate changes on the whole ecosystem.
The timing of resource availability and quality are assumed to be central in the mechanisms of population regulation of large herbivores (Mysterud et al. 2003). These latter are adapted to give birth during the vegetation green-up period in order to maximize the access to nutritious forage during high energetically demanding periods of late gestation and lactation (Rutberg 1987; Clutton-Brock et al. 1989). The access to high-quality resources after winter scarcity is key for reproductive success, rebuilding body reserves, juvenile growth and survival and finally population dynamics (Pettorelli et al. 2005b, 2007; Herfindal et al. 2006). In alpine ungulates, the protein content of the diet correlates with plant phenology during the growing season, typically rising with the increasing access to high-quality vegetation following snow melt and plant emergence (Hamel et al. 2009; Gálvez-Cerón et al. 2013). However, the ongoing increase in spring temperatures is advancing the timing of plant growth in temperate regions and therefore are also time-shifting the access to high-quality vegetation (Pettorelli et al. 2005b; Cleland et al. 2007). Because plant growing season is a key period for the growth, reproduction and survival of alpine herbivores, any factor affecting these seasonal patterns of forage quality and availability may have long-lasting effects on their life-history traits (Gaillard et al. 1996; Solberg et al. 2008). In arctic ecosystems it has been already shown that climate change disrupts population dynamics and trophic interactions (Post & Forchhammer 2008; Hansen et al. 2013). Thus, studying the dietary consequences of environmental change is fundamental to understand the evolutionary and demographic processes in ungulate populations, especially in the context of the ongoing climate change (Nielsen et al. 2012).
Climatic variability is not the only process affecting the diet of large herbivores, and ultimately population dynamics. Population density has also a clear impact on resource availability due to intraspecific competition (Bonenfant et al. 2009). High densities alter the habitat use (Mobaek et al. 2009), shape the diet composition (Kausrud et al. 2006) and therefore affect the diet quality (Blanchard et al. 2003). Furthermore, density-dependent processes often interact with climatic variations in a complex interplay (Bonenfant et al. 2009). Usually, intraspecific competition is stronger when climatic conditions (and therefore resource abundance) are unfavorable and harsh climatic conditions impose sharper constraints in populations experiencing high densities (Coulson et al. 2001; Fattorini et al. 2018a).
While climatic and demographic fluctuations should influence the diet of large herbivores through resource availability and quality, individual traits should furthermore influence resource selection processes through, for instance, age- and sex-specific foraging behavior and habitat selectivity (Marchand et al. 2015). Increases in foraging efficiency with age and experience have been documented in several vertebrates, especially birds (Daunt et al. 2007). Although much less commonly reported, a new body of evidence reveals patterns of increased experience and intraspecific aggression over life in ungulates (Fattorini et al. 2018b; Froy et al. 2018), potentially resulting in higher protein intake at old ages. Likewise, female large herbivores are constrained to compensate for the increased energetic requirements linked to pregnancy and lactation and should have a diet higher in protein than males during the growing season (Miranda et al. 2012).
So far, the effect of density-dependent and density-independent factors on the use of resources by large herbivores have most often been studied by combining rough proxies of food availability and quality, such as habitat categories, vegetation indexes and meteorological variables (Pettorelli et al. 2005a; Nielsen et al. 2012; but see Duparc et al. 2020). This is often due to logistical constraints preventing the collection of dietary information on individuals. Accordingly, very few studies have been able to link climate change, population density, individual characteristics and diet quality, through individual foraging choices in term of diet composition. We aimed at filling this gap by studying the effect of global warming on spring onset and its consequences on the dietary composition and quality of a medium-size herbivore, considering density-dependent processes and individual characteristics. We benefited from an exceptional long-term dietary monitoring (24 years) of a Pyrenean chamois population (Rupicapra pyrenaica pyrenaica Bonaparte, 1845), a mountain ungulate highly selective for high-quality plants (Duparc et al. 2020), experiencing large variation in population density (Beaunée et al. 2015) and facing an earlier onset of spring (see Results and Fig. 1a). Working with a direct individually-based measure of diet quality (fecal nitrogen; Gálvez-Cerón et al. 2013), we monitored >250 individuals and were provided with a unique opportunity to assess consistent mechanistic links between environmental changes and dietary consequences, while accounting for age- and sex-specific effects on foraging behavior.

More specifically, we first studied the relationship between diet quality and diet composition in order to understand which type of plants enhance the dietary quality of this highly selective herbivore. We then hypothesized that an earlier onset of spring will advance the access to more nutritious and digestible plants, increasing their consumption (Menzel & Fabian 1999) and the diet quality (as indexed by fecal nitrogen) of chamois. Conversely, high population abundance should force individuals to feed on resources with lower nutritional quality due to intra-specific competition (Stewart et al. 2011). This effect could interact with variation in spring onset, increasing in strength in years with later springs (Bonenfant et al. 2009). We also expected these patterns of diet composition and quality to be influenced by selection processes related to individual characteristics, with females expected to select for higher quality food items than males due to their increased energetic requirements during spring (e.g. Miranda et al. 2012). Similarly, old individuals might benefit from their experience in selecting and securing the best food resources in term of quality (Fattorini et al. 2018b; Froy et al. 2018).
MATERIALS AND METHODS
Study area and chamois data collection
The study was carried out in the National Game and Wildlife Reserve of Orlu (NGWR-O), Eastern Pyrenees, France (42.66°N, 1.97°E). The reserve is an area of 4243 ha ranging from 950 to 2765 m.a.s.l. extending from montane to nival ecosystems. The area is characterized by a mountainous oceanic climate with annual mean temperatures of 9.2°C and mean yearly-accumulated rainfall of 1260 mm. The mountain stage is mainly composed of beech forests (Fagus sylvatica) with a substrate of genus Vaccinium, Galium and Luzula among others. The subalpine stage is mainly composed of heathlands dominated by genus Rhododendrum and Vaccinium, grasslands of Festuca spp. and Nardus stricta and scarce mountain pine (Pinus uncinata) forests. Finally, grasslands, rocks and cliffs are the prevailing habitats in the highest slopes.
The abundance of Pyrenean chamois in the NGWR-O has been monitored since 1984. Abundance was initially monitored by annual single block counts in late spring from 1984 to 2008 and was progressively replaced in 2005 by helicopter surveys replicated 5 to 8 times each year to determine uncertainty around abundance estimates. Aerial counts were numerically scaled to block counts (see Gilot-Fromont et al. 2018). Although these approaches systematically underestimate the true population size to a variable extent (Morellet et al. 2007), they provide a relatively fair overview of the long-term trend in population abundance due to the very contrasted demographic periods encountered (Beaunée et al. 2015). Indeed, since the early 1990s, this population has been repeatedly infected by Border Disease virus, a pestivirus that has caused dramatic decreases, and in some cases collapses, of Pyrenean chamois populations during the last 30 years along the Pyrenees (Marco et al. 2015; Lambert et al. 2018). As a consequence, the abundance of chamois in the NGWR-O has continuously decreased since the first contact with the virus and until the early 2010s when the virus disappeared from the population without intervention (Gilot-Fromont et al. 2018). In this population, most births are highly synchronized and occur between mid-May and mid-June, with a peak during the first week of June (Loison 1995).
Concurrently, a long-term capture-mark-recapture monitoring has been performed since 1985 (Loison et al. 1999). The number of animals trapped each year varied from 2 to 66, using different methods such as corral nets, snares, drive nets or darting. The sex and age of animals were recorded since 1985 and fecal samples were collected on captured animals since 1992 (except during 1994 and from 1996 to 1999). It should be noted that fecal pellets are mainly representative of the 24 h previous diet (Espunyes et al. 2019b). The age was determined by counting horn growth annuli (Corlatti et al. 2015) and fecal droppings were collected and stored frozen at −20°C until laboratory analysis. The captures and sampling of Pyrenean chamois were performed by the Office Français de la Biodiversité, under the authority of the Ministry of Agriculture, Agrifood, and Forestry and the Ministry for the Ecological Transition.
Other herbivores such as red deer (Cervus elaphus Linnaeus, 1758) and roe deer [Capreolus capreolus (Linnaeus, 1758)] are present at low densities in the study area. A herd of 1600 domestic sheep (Ovis aries)—whose number has not significantly changed during our study period— is also present during the summer months. This herd has been present since several decades before the start of our sampling. A negligible effect of inter-specific competition for food resources is thus expected for chamois during spring period (April-June; see below).
Plant phenology

Diet quality
All fecal samples collected on captured animals in April-June from 1992 to 2015 were analyzed (n = 277, median date of capture: April 28). Frozen feces were thawed and oven-dried at 60°C for 24 h and then ground with a 1-mm pitch laboratory mill (Cyclotec 1093, FOSS Tecator, Höganäs, Sweden).
The fecal nitrogen (fN), acid-detergent fiber (fADF) and lignin (fL) estimations were carried out using Near-Infrared Spectroscopy (NIRS), a method already calibrated for fecal samples of Pyrenean chamois (Villamuelas et al. 2017). The fN is a reliable indicator of diet quality widely used in studies of herbivores diet (Robbins 1983; Hamel et al. 2009) while fL, as a digestive retardant, is a complementary indicator of low digestibility and low nutritive content of forage (Hatfield & Fukushima 2005). The fADF was used to correct fN values following the recommendations of Gálvez-Cerón et al. (2015). Ground fecal samples were scanned over a wavelength from 1100 to 2500 nm using a NIRSystems 5000 scanning monochromator (FOSS, Hillerød, Denmark). Reflectance was recorded at 2 nm intervals and each sample was scanned in duplicate. Finally, WinISI III software (v. 1.6) was used to analyze the spectra and predict fN, fADF and fL values.
Diet composition
We performed a fecal cuticle microhistological analysis on a subset (n = 155) of the previous samples and obtained a representative summary of the different plant taxa ingested by each animal (Storr 1961). Up to 6 samples (when available) were randomly selected each month.
Between 2 to 3 g of each fecal sample was mixed with 3 mL of 65% concentrated HNO3. These test-tubes were placed for 1 min in a bath of boiling water and diluted with 200 mL of water. This suspension was then passed through 1.00 mm and 0.25 mm filters. Later, 50% aqueous solution of glycerine was dispersed on a microscope slide. The fraction between 0.25 to 1.00 mm was then spread on these glass slides trying to avoid any fragment overlap. Finally, cover-slips (24 × 60 mm) were fixed to the slides with DPX microhistological varnish and left to dry overnight. For each sample, 3 slides were prepared and examined by the same observer (SC) under an optical microscope at 100× and 400× magnification by viewing 2-mm- wide and 60-mm- long traverses, with 3 mm between traverses. Up to 200 fragments per sample were identified to the genus level based on the shape of epidermal cells, trichomes and stomata surrounding cells. The counting data of each plant genus were converted to percentages. Epidermal fragments of floral parts, stems, petioles, and midribs were not registered due to the difficulties in species identification. Recorded plants were pooled into 5 functional groups (see Espunyes et al. 2019a, b) for a similar approach), namely leguminous woody plants (hereafter LW), non-leguminous woody plants (NLW), graminoid plants (Gr), leguminous forb plants (LF) and non-leguminous forb plants (NLF).
Statistical analyses
We first analyzed the relationship between diet composition and diet quality by building predictive models using a partial least squares (PLS) multivariate regression approach (Carrascal et al. 2009). The PLS approach copes better with collinearity than linear models and can be used to explore relationships between a block of explanatory and correlated covariates and a block of response variables. The fN content was corrected by the fADF content (obtaining fNc), following Gálvez-Cerón et al. (2015). The proportions of plant functional groups (namely: LW, NLW, Gr, LF and NLF) were used as the predictor variables and fNc and fL concentrations were used as the response variables. We then investigated the influence of environmental and biological variables on the diet composition and diet quality of chamois. For diet composition, we used beta-regressions in which the responses variables were composed of the proportions (ranging from 0 to 0.9) of woody plants, graminoids and forbs in the diet of chamois. For diet quality, we used linear normal regressions in which fN was the response variable. The GDD during April, the abundance of chamois in the reserve, the sex and the age were included as additive predictor variables. The day of sampling was included to control for sampling design and for the rapid change in fN values during growing season (Fig. S1, Supporting Information). fADF was also included in diet quality models as predictor to control for the acid-detergent fiber content and for making fN a better proxy of diet quality (Gálvez-Cerón et al. 2015). The interaction between the abundance of chamois in the reserve and the GDD during April was also tested (Bonenfant et al. 2009). We standardized all predictor variables (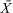 = 0, SD = 1) but sex (the only 2 levels factor), so that coefficient estimates could be interpreted as the relative contribution of each variable to the model (Gelman & Hill 2006).
= 0, SD = 1) but sex (the only 2 levels factor), so that coefficient estimates could be interpreted as the relative contribution of each variable to the model (Gelman & Hill 2006).
We tested all combinations of predictor variables. The parameter estimates for each variable included in a confidence set of models with model weight ωi > 0.05 were obtained using conditional model-averaging in the AICc framework (Burnham & Anderson 2002). Inferences based on model-averaging are recommended over model selection because the averaged estimators present a better precision and reduced bias when no single model has a clearly superior weight to the others in the set (Burnham & Anderson 2002). We checked for multi-collinearity of the predictors by using the variance inflation factors (VIF). Following (Zuur et al. 2010), if a predictor showed a VIF > 3, it would have been removed from a given model due to collinearity with other predictors. In preliminary analyses, we used Generalized Additive Models (GAM) to explore potential non-linear relationships between our predictors and fN. A non-linear relationship would have justified the incorporation of a superior polynomial term but this was not the case here, excepted for the day of sampling (Fig. S1, Supporting Information). However, for this covariate, the relationship only detracted from a linear pattern in the very last days of June and including a quadratic relationship in the best model selected (see Table S3, Supporting Information) for this covariate did not change the outcome (results not shown). We thus only kept a linear effect of the day of sampling in our models.
Because the response variables and most of our predictors presented a trend over time (see Results, Fig. 1, Fig. S2 and Table S1, Supporting Information, for an overview of temporal variation in the tested parameters), we were confronted with an inflated probability of detecting spurious correlations and confounding effects (Graham 2003). Therefore, we also detrended these predictors (GDD, abundance of chamois, sampling date, age and fADF) by regressing them on year and calculating the residuals (see Grosbois et al. 2008 for a detailed explanation). Again, we also standardized these variables and used beta/linear regressions to assess the consistency of the previous model averaging using the detrended variables in place of the raw covariates and by including the linear effect of year in all models to account for time trends in the variables. The results can be found in Supporting Information S1 and Table S2.
All statistical analyses were performed using R software version 3.6.3 (R Core Team 2020) and the accepted significance level was fixed at P < 0.05. The R-packages “mgcv” (version 1.8-23; Wood 2006) and “plsdepot” (version 0.1.17; Sanchez 2012) were used to create the variable FNc and the PLS multivariate regression between diet composition and quality, respectively. The R-package “MuMIn” (version 1.43.6; Barton 2018) was used for model averaging and the package “car” (version 3.0-0; Fox & Weisberg 2011) was used to calculate the VIF of the variables. The package “betareg” (version 3.1-3; Cribari-Neto & Zeileis 2010) was used to perform the beta regressions and the package “visreg” (version 2.5-1; Breheny & Burchett 2017) was used to represent the effect of a predictor variable of interest on the response variable after correcting for the effect of the other model predictor variables (partial residual plots).
RESULTS
Chamois diet
In all the studied period (1992–2015; no data in 1994 and 1996–1999), 39 plant genera were detected in Pyrenean chamois feces samples collected during spring, but none of these genera were detected in all the samples. Overall, the most consumed group was graminoids (Gr; 40.5%, CI95 = 36.9–44.2), mainly composed of Festuca spp. (26.1%, CI95 = 23.3–28.9) and Carex spp. (7.2%, CI95 = 6.2–8.3), followed by the non-leguminous woody plant group (NLW; 25.3%, CI95 = 22.1–28.6) dominated by Calluna vulgaris (8.7%, CI95 = 6.6–10.8) and Rhododendron ferrugineum (6.5%, CI95 = 5.0–8.0). Non-leguminous forbs (NLF; 15.6%, CI95 = 13.1–18.1), leguminous forbs (LF; 9.1%, CI95 = 7.3–11.0) and leguminous woody (LW; 8.3%, CI95 = 6.4–10.2) comprised the rest of the diet, mainly represented by the LW Cytisus spp. (8.2%, CI95 = 6.3–10.0) and the LF Trifolium spp. (5.2%, CI95 = 5.0–6.5).
Relationship between diet quality and diet composition
The PLS multivariate regression was built using 2 latent vectors that explained 72.9% of the variance in diet composition and 45.6% of the variance in diet quality. The examination of the 2-dimensional regression coefficients showed that fecal nitrogen was positively related to forbs consumption (LF: 0.241, NLF: 0.281) and negatively related to graminoids (−0.141) and woody plants (LW: −0.119; NLW: −0.151). At the same time, fecal lignin was positively related to woody plant consumption (LW: 0.190; NLW: 0.283) and negatively related to graminoids (−0.453).
Effects of environmental and biological parameters on diet composition and quality
Model-averaging provided strong support for an effect of sampling day and abundance on all components of the diet composition and quality of Pyrenean chamois (Table 1). Similarly, the importance of GDD in explaining variation of fN over time was highly supported, whereas this covariate had no effect on diet composition. Among individual characteristics, only age had an effect on graminoid consumption and diet quality. Models including other factors (sex and interaction between abundance and GDD) had much less support and high uncertainties around their model-averaged parameters estimates. For each response variable, we considered the model including significant model-averaged parameter estimates as the best model to compute the variance inflation factors. For every variables included in the best models, none of the VIF computed were >2, providing no evidence for collinearities among predictors (Table S4, Supporting Information).
| Parameter | Response variable | Relative importance | Averaged estimate | SE | z-value | P-value |
|---|---|---|---|---|---|---|
| GDD | fN | 1 | 0.139 | 0.039 | 3.52 | <0.001 |
| Woody plants | 0.30 | −0.006 | 0.096 | 0.06 | 0.94 | |
| Graminoids | 0.47 | −0.013 | 0.105 | 0.13 | 0.89 | |
| Forbs | 0.35 | 0.011 | 0.091 | 0.125 | 0.90 | |
| Abundance | fN | 1 | −0.145 | 0.038 | 3.76 | <0.001 |
| Woody plants | 1 | 0.418 | 0.093 | 4.49 | <0.001 | |
| Graminoids | 1 | −0.270 | 0.097 | 2.77 | 0.005 | |
| Forbs | 1 | −0.265 | 0.091 | 2.90 | 0.003 | |
| DoY | fN | 1 | 0.366 | 0.033 | 10.89 | <0.001 |
| Woody plants | 1 | −0.319 | 0.091 | 3.49 | <0.001 | |
| Graminoids | 1 | −0.247 | 0.085 | 2.88 | <0.001 | |
| Forbs | 1 | 0.60 | 0.07 | 7.56 | <0.001 | |
| Age | fN | 0.85 | 0.064 | 0.029 | 2.19 | 0.027 |
| Woody plants | 0.44 | 0.101 | 0.077 | 1.31 | 0.19 | |
| Graminoids | 0.82 | −0.173 | 0.074 | 2.33 | 0.019 | |
| Forbs | 0.28 | 0.045 | 0.068 | 0.66 | 0.51 | |
| Sex | fN | 0.30 | 0.062 | 0.066 | 0.94 | 0.35 |
| Woody plants | 0.23 | 0.056 | 0.173 | 0.32 | 0.75 | |
| Graminoids | 0.23 | 0.035 | 0.173 | 0.203 | 0.83 | |
| Forbs | 0.23 | 0.001 | 0.142 | 0.01 | 0.99 | |
| fADF | fN | 1 | −0.194 | 0.029 | 6.63 | <0.001 |
| Abundance × GDD | fN | 0.35 | −0.040 | 0.042 | 0.95 | 0.34 |
| Woody plants | 0.09 | −0.097 | 0.117 | 0.83 | 0.40 | |
| Graminoids | 0.23 | 0.161 | 0.112 | 1.43 | 0.15 | |
| Forbs | 0.12 | −0.104 | 0.114 | 0.91 | 0.36 |
- † Relative importance represents the sum of the AICc weights across all the models where the predictor occurs. GDD, the accumulated growing degree during April; DoY, the day of sampling; fADF, the fecal acid-detergent fiber content
Model-averaged parameter estimates indicated that the consumptions of woody plants and graminoids were negatively related to sampling day. On the contrary, the consumption of forbs and diet quality was positively related to this parameter.
Both GDD (slope ± SE = 2.15 ± 0.93, R² = 0.19, P = 0.030) and abundance of chamois (slope ± SE = −47.19 ± 3.92, R² = 0.87, P < 0.001) showed trends over time, characterized by earlier onsets of spring and decreasing estimates of population size since 1992 (Fig. 1). These shifts contributed to changes in diet composition and quality (with similar effect size of both factors for models on diet quality; Table 1). Decreasing abundance of Pyrenean chamois was associated with a decreasing consumption of woody plants (−56% between extreme abundance values; Fig. 2) and a concomitant increase in diet quality (Fig. 3). By contrast, the consumption of graminoids and forbs increased with decreasing abundance of chamois (+87% and +143% between extreme abundance values, respectively). Increasing values of GDD in April, and the related advance of spring onsets, also resulted in a marked increase in fecal nitrogen (+22% for the most extreme values; Fig. 3) without any effect on diet composition (Table 1). Finally, old animals of both sexes had a diet composed of less graminoid than young animals, and of higher quality (Fig. 3).


DISCUSSION
Our long-term study shows that the ongoing advancement of spring onset (Fig. 1a) is leading to an earlier access to highly nutritious early-growing plant stages, increasing diet quality in Pyrenean chamois. However, these earlier spring onsets had no effect on the diet composition, this latter being rather related to population densities. Thus, while consuming the same proportions of plant groups, chamois obtained a more nutritious diet during earlier springs.
Herbivore species living in highly seasonal environments are expected to make foraging choices that track the timing of spring vegetation growth and the seasonal availability in high-quality plants (Post & Stenseth 1999; Zweifel-Schielly et al. 2012; Espunyes et al. 2019a; Duparc et al. 2020; see also Fig. S1, Supporting Information). This is particularly expected in medium-sized ungulates like chamois in which species’ energy requirements and digestive abilities make them more selective for high-quality food (Clauss et al. 2003). In our study, we observed that Pyrenean chamois increased their consumption of forbs throughout spring but decreased the consumption of graminoids and woody plants. This feeding process is directly related to the pattern of dietary quality as indexed by fecal nitrogen. FN is considered in many studies as a reliable indicator of phenology and quality of plants eaten by grazing and mixed-feeding herbivores (Festa-Bianchet 1988; Hodgman et al. 1996). Accordingly, our results show an increase of fecal nitrogen content along the season related to an increasing consumption of forbs and a decreasing consumption of graminoids and woody plants. In highly seasonal environments, growing plants present seasonal changes in nitrogen concentrations. Deciduous plants allocate substantial quantities of nitrogen into new leaves during early growing stages (i.e. spring; van Soest 1994). However, evergreen plants allocate it gradually throughout the growing season and in lower quantities, making deciduous plants more appetizing (Jaeger et al. 1999; Körner 2011). In fact, in our study, the consumption of forbs throughout spring varies much more than the other plant groups, supporting the importance of its phenology in its consumption. Similar findings have been reported in an Alpine population of chamois where animals systematically selected a low proportion of the available biomass composed of high-quality plants at a specific phenological stage (flowering) throughout the growing season (Duparc et al. 2020).
We detected an earlier onset of the growing-season over the last decades (Fig. 1a) in line with several studies that provide evidences about climate warming and advances in the phenological traits of organisms (Menzel & Fabian 1999; Parmesan 2006; Moyes et al. 2011). By allowing individuals more time to grow and build up reserves, this phenomenon could lead to an enhancement of the performance of herbivores, as reported in other populations and species (Herfindal et al. 2006; Ozgul et al. 2010; Garel et al. 2011; Tveraa et al. 2013). However, this is not always the case as Pettorelli et al. (2007) determined that warmer springs also tended to be associated with fast plant growth and shortened periods of high quality forage, that in turn negatively affected juveniles growth in 2 alpine ungulate species. Similar counterproductive effects may occur in the growth and survival of juveniles if the species is unable to adjust its parturition time to the earlier springs, causing a mismatch between parturition and the optimal foraging period (Post & Forchhammer 2008). Plard et al. (2014) detected that this mismatch led to decreasing dynamics in a roe deer population. Kourkgy et al. (2016) already questioned the ability of Pyrenean chamois to adjust their gestation length or conception dates to the earlier onsets of spring. Their results showed that females had a much lower adaptive potential to the spring conditions than to autumn conditions. However, capital breeders, as Pyrenean chamois, should meet the high cost of early lactation mainly through maternal capital, potentially reducing the adverse effects of this mismatch (Kerby & Post 2013; Richard et al. 2017). Furthermore, the impact of climate warming during the seasons generally considered as most important (spring and winter in alpine/artic ecosystems) could also be counterbalanced by other consequences in other seasons traditionally considered as less important (e.g. autumn: Loe et al. 2020). In that sense, more research on the demographic impacts of phenological mismatch and other consequences of climate warming over seasons in alpine species is needed to determine the consequences of climate change over the long term.
Our findings also show that increasing population density negatively affects individual diet quality. As animal density increases, it is expected that intraspecific competition for resources also increases, affecting the access to the most nutritious plants at the individual level (Kie et al. 1980; van Beest et al. 2014). It is thus fundamental to consider this parameter when evaluating the mechanisms and the consequences of global warming on the life-history traits of herbivores. Additionally, the abundance of chamois in the area was negatively related with the consumption of forbs and graminoids, while positively related to the consumption of woody plants. In fact, the consumption of woody plants was mainly driven by the abundance of chamois and the effect was 2 and 4 times stronger than for the consumption of graminoids and forbs, respectively. Lignin is not only indigestible, but also interferes with digestion by reducing the rumen turnover rate and the intake capacity (Van Soest 1994). Thus, the consumption of graminoids, while negatively related with nitrogen intake, can be favorable due to their highly negative relationship with fecal lignin content. Therefore, intraspecific competition forces chamois to have a lower digestible and protein-rich diet and, conversely, the important negative trend over time in chamois abundance has led individuals of this population to feed much less on woody plant and more on graminoids and forbs. Note that this pattern could have been also generated by temporal changes in landscape characteristics (e.g. open-landscape restoration; Cazau et al. 2011). However, this is not the case here and, instead, the proportion of grasslands over woody habitats has rather decreased by around 12% during the study period —a common process in alpine areas (Venn et al. 2014)—making our results conservative (see Supporting Information S2).
Older animals had a higher diet quality than younger ones and consumed lower proportions of graminoids. These results could provide additional support to the theory of an increased experience in plant selection for herbivores during their life (Froy et al. 2018; Malagnino et al. 2021) but could also be explained by a dominance hierarchy (Fattorini et al. 2018b). However, contrary to the predictions, sex had no effect on the diet quality and composition, despite a sexual difference in the spatial behavior and habitat selection of this species (Loison et al. 1999; Dalmau et al. 2013). Sexual differences in diet quality and composition has rarely been studied in any chamois, but Pérez-Barberia et al. (1997) observed a similar diet composition in both sexes, at least for Cantabrian chamois (Rupicapra p. parva) in spring. Being highly selective for high-quality palatable plants (Corlatti et al. 2013; Redjadj et al. 2014; Duparc et al. 2020) chamois species may be able to feed selectively on high-quality forage and compensate for sex-specific behavioral differences.
We recommend the use of fecal nitrogen to monitor long-term trends in density-dependent and density-independent environmental conditions through changes in the nutritional state of a given population (Blanchard et al. 2003; Gálvez-Cerón et al. 2013). Furthermore, because we did not detect a sex effect and age has less effect (2.3 times lower) as compared to densities, the collection of fresh unidentified droppings during seasonal field surveys coupled with NIRS analysis could be an inexpensive option to monitor the density-dependent processes in a population as a complement to other indicators of ecological changes (Morellet et al. 2007; Santos et al. 2014). This study also highlights the fundamental need of long-term monitoring programs to understand the ecological processes affecting herbivore performance in the context of global changes (Festa-Bianchet et al. 2017).
In the current context of climate crisis (Steffen et al. 2018), understanding the mechanistic effects of these variations on the dietary selection of individuals is an essential step to understand the consequences of climate change on the life-history traits of wild populations (Sinclair & Parkes 2008). Projection studies on species distribution, food webs and trophic asynchrony are in crucial need of data on the ecological mechanisms of environmental change and associated dietary selection (Thuiller et al. 2017; Veldhuis et al. 2019). By including data on resource use and dietary quality, we believe that such studies may create more realistic and robust predictions on the impact of the climatic crisis on the life-history traits of wild herbivores.
ACKNOWLEDGMENTS
The authors are thankful to all the field assistants and volunteers for their amazing work in trapping Pyrenean chamois and collecting feces during such a long period. They are grateful to Météo France for providing the climate data. This study was funded by the research partnership program “Approche Intégrée de la Démographie des Populations d'Isard” (N°2014/08/6171) between the Office Français de la Biodiversité (formerly Office National de la Chasse et de la Faune Sauvage) and the Servei d'Ecopatologia de la Fauna Salvatge (SEFaS). Johan Espunyes acknowledges a pre-doctoral grant from the Government of Andorra (ATC015 AND 2015/2016, 2016/2017, and 2017/2018). Emmanuel Serrano was funded by the Spanish Ministerio de Economía y Competitividad (Mineco) through a Ramon y Cajal agreement (RYC-2016-21120).

