

Telomere Length Is Associated With Obesity Parameters but With a Gender Difference
Abstract
Cardiovascular disease (CVD) and obesity have been coupled to short telomere length in peripheral blood. The biological background to this observation is not obvious from the literature. In this study we have analyzed a large set of known risk factors for CVD in relation to telomere length in blood cells on a merged cohort of 989 individuals recruited in the Malmö Diet and Cancer Cohort (MDCC) and the Northern Sweden MONICA project. We found a significant or borderline association between obesity parameters and telomere length in women after age and center adjustments (BMI: r = −0.106, P = 0.021, weight: r = −0.087, P = 0.060, waist circumference: r = −0.099, P = 0.032, hip circumference: r = −0.128, P = 0.005). In men, a positive borderline correlation to high-density lipoprotein (HDL) (r = 0.111, P = 0.053) and a negative correlation to 2-h post-oral glucose-tolerance test (OGTT) was observed (r = −0.202, P = 0.045). In neither group any association was found between telomere length and cholesterol, serum triglycerides, serum low-density lipoprotein, plasma insulin, blood pressure, pulse pressure, or smoking habits. Our data indicate that telomere length is associated with an “obesity-phenotype” but only in women.
Introduction
Telomeres are repetitive structures at the end of mammalian chromosomes and together with associated proteins they cap and protect chromosome ends (1). In normal cells, a progressive telomere shortening accompany each cycle of replication (2) and there is a well-established gradual loss of telomere length with increasing age in human peripheral blood (3,4). Shorter telomere length in peripheral blood has been shown in patients suffering from coronary heart disease compared to controls (5,6,7,8,9), but contradictive results have also been reported (10,11). Short telomeres were also associated with decreased survival, mostly due to a higher mortality rate from heart and infectious diseases in these individuals (12). Increased oxidative stress is a characteristic feature in patients with cardiovascular disease (CVD) (13). A hallmark of atherosclerosis is internalization of oxidized low-density lipoproteins by macrophages leading to foam cell formation. In endothelial cell cultures, oxidized low-density lipoprotein reduces telomerase activity and accelerates senescence, which is in agreement with data showing that oxidative stress can induce enhanced telomere attrition and premature senescence in this cell type (14). Short telomeres in peripheral blood have also been described in diabetic patients (9,15,16,17,18) and in individuals with cardiovascular risk factors, i.e., increased pulse pressure (19), obesity, increased carotid intima media thickness, high fasting glucose levels, and smoking (9,20), as well as in response to psychosocial stress (21). Taken together, these data strongly indicate that telomere length, in vivo as well as in vitro, can serve as a biological marker for age-, and stress-related conditions.
There are two possibilities to explain these observations. First, in a tissue-specific fashion, short telomeres might indicate senescence of (stem) cells, which might contribute to age-related functional impairment in this tissue. Second, short telomeres might simply indicate a history of high stress and damage and could thus act as a risk marker for age-related disease residing in many different tissues. The second possibility is most likely for the observations made on peripheral blood cells and referred to above. Interestingly, genetically modified mice with short and dysfunctional telomeres were protected from diet-induced atherosclerosis in apolipoprotein E-null mice, indicating that telomere attrition resulting in replicative senescence may also serve as a mechanism for restricting atheroma progression (22).
The aim of this study was to investigate telomere length in relation to well-known cardiovascular risk factors. Because both telomere biology and cardiovascular risk factors could differ between men and women, we have also stratified for sex.
Methods and Procedures
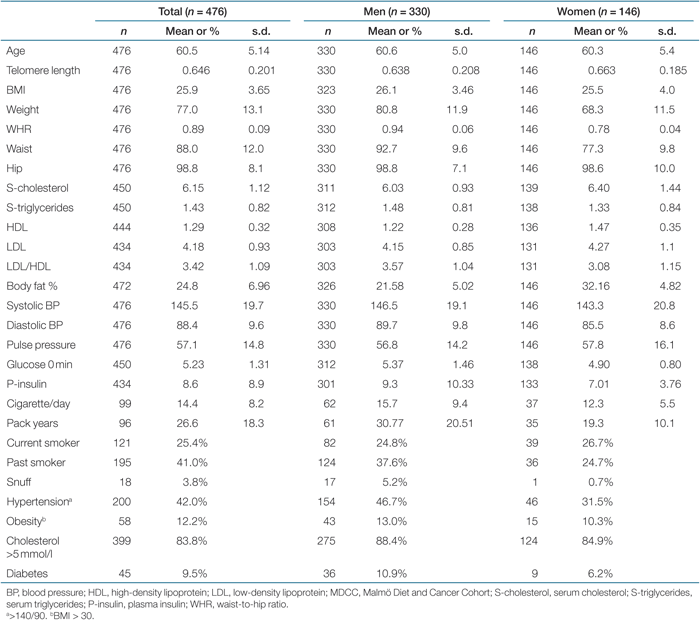
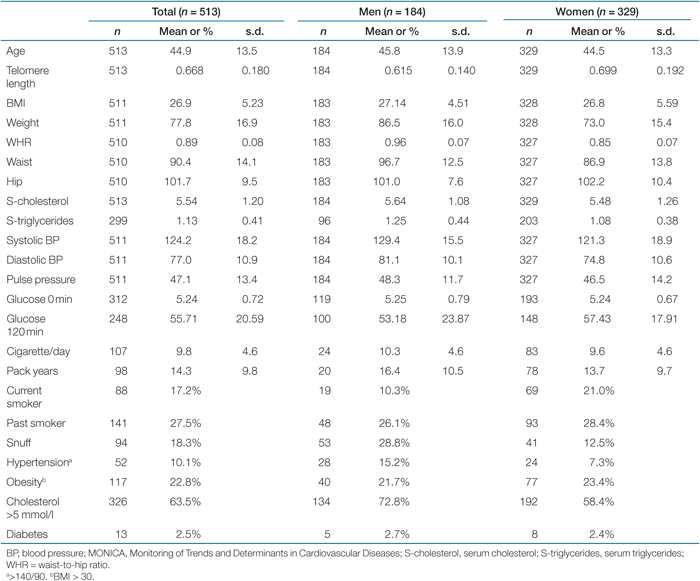
The study population consisted of 989 individuals and was a merged cohort from a part of the Malmö Diet and Cancer Cohort (MDCC) (n = 476, age range 48–68, median 62 years) and the Northern Sweden MONICA 2004 survey (n = 513, age range 26–75, median 42 years), which is a branch of the World Health Organization MONICA Project (Monitoring of Trends and Determinants in Cardiovascular Diseases). Blood was drawn and DNA extracted from granulocyte preparations and buffy coats. Serum analyses and other measurements are described elsewhere (23,24). The population-based cohorts were not screened for specific diseases but a self-administered questionnaire including demographics, medical conditions, and lifestyle characteristics was completed.
Buffy coats from individuals in the MONICA cohort were collected in 2004 and kept frozen at −80 °C. Blood samples from the MDCC study were obtained between 1992 and 1996, and the granulocyte fractions were kept frozen. DNA extraction was performed using standard procedures. Telomere length was investigated by real-time PCR (25,26) using telomere-, and control-gene (β-globin) primers in separate plates. All samples were tested in triplicates. The ΔΔCt method with a reference DNA was used for telomere length evaluation and a standard curve was run on each plate to monitor PCR efficiency. The mean interassay coefficient of variation evaluated for this assay on human blood was 3.96% when run by two investigators in the laboratory. This study was approved by the ethical committees in Malmö (#LU 51-90) and at Umeå University (Dnr 05-100M).
Statistical analyses were performed using SPSS, version 15.0 (Statistical Package for the Social Sciences, SPSS, Chicago, IL). To achieve a normal distribution, log-transformed telomere length was used in all analyses. Analyses of covariance were performed for age-, sex- and center-adjusted comparison of dichotomous variables and partial correlation for continuous, adjusted parameters. A P value <0.05 was considered to be significant.
Results
Baseline characteristics and gender differences for the MDCC and the Northern Sweden MONICA project (MONICA) are shown in Tables 1 and 2. The most prominent differences between the two cohorts were younger individuals and lower blood pressure in the MONICA project. The prevalence of smoking was higher in the MDCC study, whereas snuff was used more often in northern Sweden. To justify merging of the two cohorts, telomere length after age adjustment was compared, which showed no difference (P = 0.187). To further exclude a possible cohort discrepancy, data adjusted for center is shown. However, this did not critically change the statistics.
 |
 |
As expected, there was a negative correlation between telomere length and age (r = −0.119, P < 0.001) (Table 3). The telomere loss per year was significant for women, but not for men (P < 0.001 and P = 0.956, respectively). Women showed overall longer telomeres than men (P < 0.001) (data not shown).
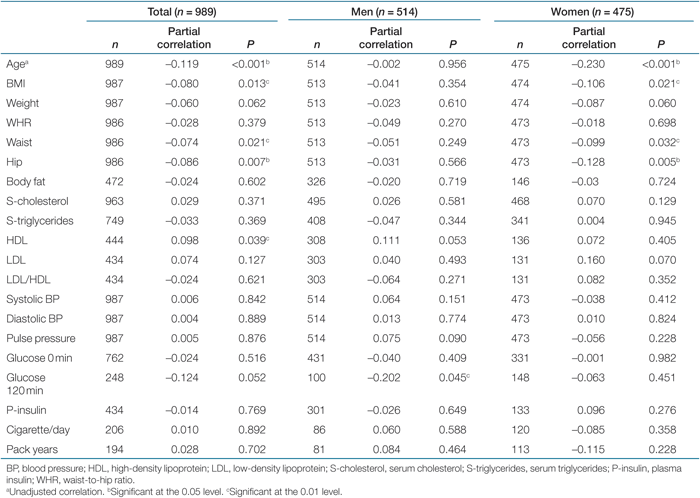 |
When analyzing telomere length in association with continuous variables after age, sex and center adjustments, a negative significant or borderline relationship was observed between telomeres and obesity parameters (BMI: r = −0.080, P = 0.001, weight: r = −0.060, P = 0.062, waist circumference: r = −0.074, P = 0.021, hip circumference: r = −0.086, P = 0.007). A positive correlation to high-density lipoprotein (HDL) (r = 0.098, P = 0.039) and a negative correlation of borderline significance to 120-min post-oral glucose-tolerance test (OGTT) was also found (r = −0.124, P = 0.052) (Table 3). To investigate possible gender effects, the analysis was performed separately in men and women. Interestingly, the telomere length was correlated with obesity parameters only in women, and to HDL and OGTT only in men (Table 3). Two age groups were created using mean age (52 years) as a cutoff. The negative association to obesity parameters was stronger, although not significant for all parameters, in the younger group compared to the older group. Moreover, in the older group, the difference between genders was not as marked as in the younger individuals (data not shown).
When dichotomizing the CVD risk factors at either mean value or clinically accepted cutoff levels the same pattern was observed. When comparing individuals with BMI ≤25 kg/m2 and BMI ≥30 kg/m2, a significant difference in telomere length was found (overall: P = 0.045; women: P = 0.077, men: P = 0.419, data not shown).
We could, however, not support the previously suggested association between short telomeres and smoking, hypercholesterolemia, plasma insulin, diabetes, hypertension, and pulse pressure (Table 4).
 |
Both inflammation and oxidative stress have been shown to influence telomere length (9,14). Self-reported rheumatoid-like joint and muscle problems were the only diseases recorded in the MONICA study that resembled those conditions. However, individuals who reported joint and muscle problems did not differ with respect to telomere length compared to the rest of the cohort (n = 49, P = 0.449 and n = 41, P = 0.164, respectively).
Discussion
In this study encompassing 989 middle-aged individuals, we found a significant or borderline correlation between telomere length and obesity parameters, but surprisingly only in women. Furthermore, a positive borderline correlation to HDL and negative correlation to postload glucose levels was observed in men. However, HDL and OGTT data were not available on all patients, and because of the low number of individuals examined and the borderline significance, these results should be interpreted with caution.
Unexpectedly, the well-established yearly telomere loss was not observed for men in the present cohort. One reason could be that older individuals in poor health were less willing to participate in studies because of their poor condition. Because of this, old individuals with presumably short telomeres could be missing in our study population. It has also been shown that men have a lower study attendance than women (27). This could provide an explanation to our puzzling observation. However, women had longer telomeres than men in our cohort, which is in agreement with previously published results (28,29,30).
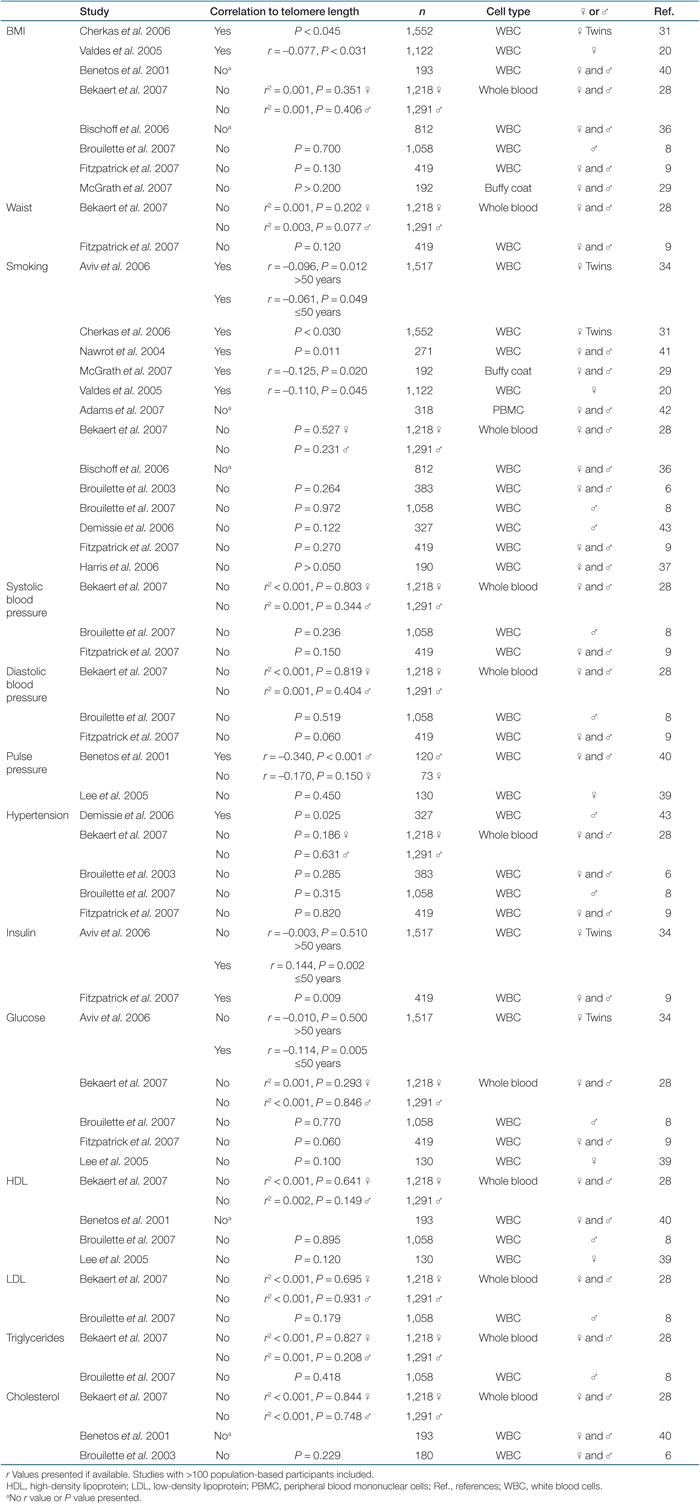
Few studies on telomere length and CVD risk factors have investigated men and women separately, but our data indicate that stratifying for gender when performing statistical calculations are important. There exists an inconsistency in the literature regarding telomere length and its association with BMI but interestingly, the studies including only women (20,31) have shown a significant negative correlation to BMI whereas the one study including only men (8) did not, giving strong support to the data presented here (Table 4). High waist-to-hip ratio (i.e., greater waist circumference than hip circumference) is normally considered as a risk factor for CVD. The negative association found between telomere length and both hip and waist circumference and the lack of correlation to waist-to-hip ratio in this study is therefore interesting. This implies a body shape effect independent of the fat distribution symmetry. We cannot rule out a possible colinearity effect among parameters of the metabolic syndrome. Insulin resistance, lipids (including low HDL), and other CVD risk factors are intimately associated with metabolic disturbances and thus have a relationship with obesity (32). Therefore, the different findings for men and women in this study can both be coupled to obesity, giving short telomeres.
No association with smoking, blood pressure/pulse pressure/hypertension, or serum lipids, except for a positive correlation with HDL cholesterol in men, was observed. This is in agreement with some published studies, whereas others did find a correlation with these parameters (Table 4). The prevalence of smoking and the number of cigarettes smoked per day in Sweden is low compared to other countries, providing a possible explanation for the discrepancies against other publications (33). However, our study is not the only one showing poor correlation between telomere length and smoking (Table 4).
In the recently published ASKLEPIOS study by Bekaert et al., no association between telomere length and classical CVD risk factors were found (28). Instead, short telomeres were significantly correlated with an unhealthy lifestyle and several inflammation and oxidative stress markers. Therefore, the authors suggested that the link between short telomeres and CVD is due to increased cell turnover in response to the inflammation component of the disease process (28). Other publications have also observed a negative association between telomere length and C-reactive protein and interleukin-6 levels (9,34). Obesity is associated with systemic chronic inflammation (35), which also can affect telomere length by increased proliferation. Although no inflammation or oxidative stress markers were available on individuals in our cohort, the data from the ASKLEPIOS study supports the lack of association with CVD risk factors found in our study, at least for women. The only accessible data on diseases that could resemble chronic inflammation in our cohort was rheumatoid-like joint and muscle problems. As there was no difference in telomere length between individuals who report such symptoms and who do not, this aspect was not considered when performing the analyses.
The previously observed association between short telomeres and myocardial infarction seems to be coupled to individuals who develop disease at a relatively young age (8,9). These patients might be hypersensitive, or have a pathological ability of risk factor accumulation compared to older patients, causing disease at an earlier age. This risk factor hypersensitivity might give increased inflammation and oxidative stress leading to significantly abbreviated telomeres, a scenario not observable at older ages. The hypothesis is also supported by the decreased survival found in individuals with short telomeres between 60 and 74 years of age but not among those >74 (ref. 12). In our cohort, there was a stronger association between short telomeres and obesity parameters in individuals younger than mean age. In addition, a greater difference between genders could be observed in the younger group. Controversial studies showing that telomere length can predict survival have been published (10,36,37) and some of them do not show an association with survival in the oldest individuals giving further support to our finding (10,36). There might be other factors outplaying the importance of telomere length in this respect in the oldest individuals.
Telomerase is an enzyme that is able to lengthen telomeres. Its importance in CVD has been less discussed than telomere length. However, low telomerase activity has been associated with CVD risk factors and the metabolic syndrome in women (38).
The individuals examined in this study were from two cohorts showing different age distributions. In separate analyses, the two cohorts showed somewhat different results with few significant findings. It is more likely that the differences found between the cohorts were due to decreased power and a normal variation between younger and older age individuals, rather than a true cohort discrepancy. However, adjustment for center was performed to compensate for eventual cohort biases.
When investigating the possible link between telomere length and diabetes risk factors such as plasma-glucose and plasma-insulin levels, only a significant association to post-OGTT was found in men. With our present knowledge, two studies analyzing telomere length in relation to insulin levels have shown a significant correlation (9,34), but no study with >100 participants did show association with glucose levels (8,9,28,39). Telomere length in diabetic patients compared to nondiabetic individuals did not differ in our cohort. This is in contrast with previously published results (9,15,16,17,18).
Given that telomeres shorten during aging, it is likely that the significant association with the “obesity phenotype” in women is the accumulated result of several risk factors leading to enhanced telomere loss over time.
Due to great interindividual differences with respect to telomere length abbreviation with age, studies including many participants are required to obtain a high statistical power to uncover subtle associations. This is naturally also an explanation to why studies demonstrate divergent results as clearly indicated in Table 4. Compilations of this type are sometimes necessary to increase the awareness of different published data and thereby facilitating future consensus.
Telomere length maintenance is a complex process involving a large number of interacting factors and the actual telomere length in a cell is the net result of telomere shortening and lengthening activities. In this article, we show that CVD risk factors that were negatively correlated to telomere length differed between genders. Telomere length seems to be coupled to an “obesity phenotype” but clearly only in women. We therefore state the importance of stratifying for gender when analyzing datasets from this type of population-based studies.
Acknowledgment
This study was supported by grants from the Swedish Research Council, the Research Council for Social Sciences, the Heart and Chest Fund, King Gustaf V's and Queen Victoria's Foundation, Västerbotten and Norrbotten County Councils, the Swedish Public Health Institute, the Swedish Heart and Lung Foundation, Swedish Medical Society, and the Medical Faculties of Lund and Umeå Universities.
Disclosure
The authors declared no conflict of interest.

