

The links between morphological parameters and benthic invertebrate assemblages, and general implications for hydromorphological river management
Abstract
In the last decades, hydromorphological degradation of rivers has gained more attention in river management, stressing the importance of understanding the links between hydromorphology and aquatic assemblages. The present study investigated general patterns in the response of benthic invertebrate assemblages to single morphological features along naturally diversified Slovenian rivers. The whole gradient of local habitat quality (river habitat quality, RHQ) and habitat modification (river habitat modification, RHM) features, according to the Slovenian hydromorphological assessment method, was covered. Regional natural characteristics explained the low share of RHQ and RHM variability, indicating nonregional presence of morphological features. The analysis identified predominant flow and predominant channel substrate as the most important RHQ features. We found that in contrast to RHQ features, RHM features had low explanatory power. These results suggest a weaker response of benthic invertebrate assemblages to the physical alteration itself than to the effect that the alteration exerts on habitat quality features. Variance partitioning among three environmental variable groups revealed predominantly independent effects (69%) on benthic assemblages, mostly on account of regional natural characteristics (30%) and RHQ features (31%). As benthic invertebrate assemblages are adapted to the former natural conditions, a similar modification may result in different effects with regard to regional natural differences. Therefore, the low proportion of variability, explained by RHM features, might be the consequence of joint dataset from different regions. Our study gives general implications for river management, but in order to more clearly define the significance of particular modification features, we suggest further analysis within more homogeneously defined habitats that encompass regional natural characteristics. Copyright © 2014 John Wiley & Sons, Ltd.
Introduction
Hydromorphological degradation represents one of the most important anthropogenic pressures affecting river and stream environments (Richter et al., 1997; Raven et al., 2002; Feld, 2004; Schinegger et al., 2012). Direct modifications on rivers include channelization, weir and dam construction, disconnection of floodplains and clearance of riparian vegetation (Pedersen, 2009; Verdonschot, 2009), which diminish habitat diversity of rivers. With the increasing severity of morphological degradation, the connection between physical river habitat and aquatic assemblages is gaining increased attention (Davy-Bowker and Furse, 2006; Friberg et al., 2009a; Vaughan et al., 2009). Managing river systems often includes a variety of restoration projects, with the general aim to reestablish lost physical diversity by substrate manipulation (Muotka et al., 2002; Jähnig and Lorenz, 2008), including the introduction of large woody debris (Kail et al., 2007; Hrodey et al., 2008; Testa et al., 2011) or in-stream structures (Harrison et al., 2004) and rehabilitation of natural riparian vegetation (Allan, 2004; Death and Collier, 2009; Riley and Dodds, 2012). However, the results of these restoration projects indicate a negligible response of aquatic biota (Lepori et al., 2005; Jähnig et al., 2010; Palmer et al. 2010; Haase et al., 2012), emphasizing the need for improved understanding of the relationship between aquatic communities and morphological pressures.
In Europe, assessment of river status gained a new perspective with implementation of the Water Framework Directive (WFD, European Commission, 2000). The newly introduced term ‘hydromorphological’ signifies the importance of assessing habitat quality and modification in the ecological classification of rivers. In the last decades, several assessment methods that summarize the heterogeneous nature of riverine physical habitat have been developed (Muhar et al., 1996, 1998; Agences de l'Eau & Ministère de l'Environnement, 1998; Raven et al., 1998, 2003; LAWA, 2000; Munne et al., 2003; Feld, 2004). One of the more comprehensive methods is the UK River Habitat Survey (RHS) method (Raven et al., 1998, 2003) with a Habitat Quality Assessment score and a Habitat Modification Score. The application of the RHS method was tested in several European countries (Balestrini et al., 2004; Szoszkiewicz et al., 2006; Tavzes et al., 2006; Bona et al., 2008; Urošev et al., 2009; Raven et al., 2010), and an adapted version was developed for use in southern Europe (Buffagni and Kemp, 2002). The RHS method also served as a basis for the Slovenian hydromorphological (SIHM) assessment method development, where evaluation of different categories of river features was made with regard to their influence on the benthic invertebrate community (Tavzes and Urbanič, 2009), resulting in 33 collected morphological features. Using these, it is possible to classify the morphological status of survey sites (Tavzes and Urbanič, 2009) by the calculation of a river habitat quality (RHQ) index and a river habitat modification (RHM) index.
According to the WFD, the main focus of assessment methods is on biological quality elements, and the direct assessment of hydromorphology serves only as additional evaluation. The most prevalent biological quality element (Birk and Hering, 2006), with the longest tradition in river assessment, are the benthic invertebrates (Kolkwitz and Marsson, 1909; Sladecek, 1973). Benthic invertebrates represent a diverse group that integrates ecosystem changes over time and responds to different environmental stressors (Sandin and Hering, 2004; Sandin et al., 2004; Friberg et al., 2009b). In Europe, several benthic invertebrate-based assessment methods exist (Birk et al., 2012), but only some of them address hydromorphological impact (Lorenz et al., 2004; Ofenböck et al., 2004; Urbanič, 2014).
Stressor-specific assessment methods detect impairment (e.g. organic pollution and eutrophication), but, particularly when considering hydromorphological alteration, the direct cause of degradation remains unknown. This is reflected in a large amount of unsuccessful restoration attempts. It is of high importance to establish more precise links of morphological characteristics and their alteration to the response of aquatic communities. A large number of studies demonstrate that benthic invertebrate assemblages are influenced by the quality of habitat features (Lammert and Allan, 1999; Sandin and Johnson, 2004; Rios and Bailey, 2006; Lorion and Kennedy, 2009; Syrovátka et al., 2009) and are also good indicators of morphological degradation (Erba et al., 2006; Feld and Hering, 2007; Larsen and Ormerod, 2010). However, the relationship between benthic invertebrate assemblages and single morphological features included in applied assessment methods was rarely analysed; and most studies considered only RHS features (Erba et al., 2006; Hughes et al., 2008; Cortes et al., 2009; Dunbar et al., 2010). Indices of the SIHM method were used as a stressor gradient in the development of the Slovenian ecological status assessment and classification method using benthic invertebrates (Urbanič, 2014) and showed good explanatory power. But no analysis was performed using single morphological features. As the SIHM method is used for hydromorphological assessment of rivers, knowledge of the relationship between single morphological features included in the SIHM method and the response of benthic invertebrate assemblages could represent a starting point for river management.
Generally, river management authorities need to have an understanding of how river ecosystems respond to anthropogenic changes and how to reverse the degradation process. There is a request for management concepts to be applicable across larger scales and not specifically focused on individual rivers or sites. Our data provide an overview of Slovenian river morphological characteristics and their alterations across a wide geographical gradient (Urbanič, 2011). We tested the possibility of finding general patterns in the response of benthic invertebrate assemblages to river morphological features. However, regional (large-scale) parameters are known to influence local (small-scale) physical features and biota (Frissell et al., 1986; Poff, 1997; Sandin and Johnson, 2004). This should be considered when analysing the relationship between morphological features and benthic invertebrate assemblages. Therefore, the most important regional natural characteristics, also relevant for Slovenian typological delineation (Urbanič, 2011), were used in our study as a separate set of environmental variables. With a focus on the relation of single morphological features included in the SIHM method to the response of benthic invertebrate assemblages, the main aims of the present study are (i) to investigate the relationship between regional natural characteristics and morphological quality or morphological modification features across the Slovenian landscape, (ii) to define the habitat parameters that are most important in structuring benthic invertebrate assemblages and discuss their explanatory power in comparison with regional characteristics and (iii) to define what share of benthic invertebrate assemblage composition can be attributed to distinctive effects among environmental variable sets (regional natural characteristics, morphological quality and morphological modification).
Methods
Study area
Slovenia covers a total area of 20 273 km2 and has 4573 km of river channels within catchments larger than 10 km2. The rivers extend across three main ecoregions of the country (Alps, Dinaric Western Balkan and Pannonian lowland), which cover more than 99% of the area and the Po lowland that accounts for less than 1% of the area (Illies, 1978; Urbanič, 2008a). The wide ecological variety of the area is also a consequence of the connection between two main river basins (Urbanič, 2008b), big karst area with karst phenomena ( ams, 2004), variety of geologies and rivers of varied sizes (Urbanič, 2011). In Slovenia, the official river typology used for bioassessment (OGRS, 2009) was developed according to System B of the WFD Annex II. The four ecoregions are divided into 16 bioregions according to the predominant geology, altitude of the catchment area and division between the river basins (Urbanič, 2008b, 2011). Ecological river types are defined using bioregion and additional qualitative environmental parameters (e.g. river size, karst spring influence, periodical flooding and intermittency).
Benthic invertebrate data
Data on benthic invertebrate composition and abundance were obtained from 302 sites between the years 2002 and 2011 (Figure 1) as part of monitoring and assessment system development programmes in Slovenia (Urbanič et al., 2008). The aim of this study was to examine the relation of natural morphological features and their modification to benthic invertebrate assemblages. The selection of sites cover the gradient from natural to heavily altered morphological conditions. Only sites where morphological alteration was the presumed main stressor were included in the analyses. Sites showing a less than good ecological status for organic pollution (Slovenian Saprobic index – SIG3 < 0·6; Urbanič, 2011; BOD5 > 5·4 mg l−1) or nutrients (total phosphorus > 205 µg l−1, orthophosphate > 84 µg l−1 and nitrate > 9·5 mg l−1) were a priori excluded. Rivers were sampled during low-to-medium-discharge conditions, generally between May and September, except some large or intermittent rivers were sampled in winter because of their natural hydrological regime. The sampling procedure followed the standardized Slovenian river bioassessment protocol (OGRS, 2009; Pavlin et al., 2011). Each site was sampled on a single occasion using a 500-mm mesh-size hand net. The sample on each site consisted of 20 subsampling units, with a total sampling area of 1·25 m2 that was taken along a 100- to 250-m river stretch in proportion to the coverage of the microhabitat types (Urbanič et al., 2005). Microhabitat types were defined as a combination of substrate and flow type with at least 5% coverage. A total of 302 samples then underwent laboratory subsampling procedures, and benthic organisms from a quarter of each whole field sample were identified and enumerated (Petkovska and Urbanič, 2010). Benthic invertebrates were determined to the taxonomic level used for the assessment of ecological river status in Slovenia (OGRS, 2009), i.e. mostly to species level.

Environmental data
For each site, data on eight regional natural characteristics were obtained from the Slovenian river typology (typology variables – Table 1). Sites were classified into four ecoregion variables (Alps – ER4; Dinaric Western Balkan – ER5; Pannonian lowland – ER11; and Po lowland – ER3). However, we only had a few sampling sites in ER3. Because typological and natural morphological parameters are similar among ER3 and ER11, these sites were combined into a united region, lowland (ER3.11). Altitude and slope were calculated using a digital elevation model with 5-m accuracy. Each site was also classified into one of four river size classes determined by catchment area and mean annual discharge (1 – catchment size 10–100 km2; 2 – catchment size 100–1000 km2; 3 – catchment size 1000–2500 km2 and mean annual discharge < 50 m3 s−1; 4 – catchment size >2500 km2 or mean annual discharge > 50 m3 s−1).
| Environmental variable | Abbreviation | Unit | Variable group | Mediana (min–max) | Occurrence frequency (%) |
|---|---|---|---|---|---|
| Region: lowland | ER3.11 | Dummy | Typology | 80 | 26 |
| Ecoregion: Alps | ER4 | Dummy | Typology | 93 | 31 |
| Ecoregion: Dinaric Western Balkan | ER5 | Dummy | Typology | 129 | 43 |
| River size class | Size_cl | Classified 1–4 | Typology | 2 (1–4) | 100 |
| Karst spring influence | Kspring | Dummy | Typology | 60 | 20 |
| Intermittency | Intermit | Dummy | Typology | 15 | 5 |
| Altitude | Alt | m a.s.l. | Typology | 283 (1–896) | 100 |
| Slope | Slope | % | Typology | 4·4 (0–261) | 98 |
| Predominant natural bank material | bnm | Score totalb | RHQ | 16 (0–44) | 96 |
| Bank features | bf | Score totalb | RHQ | 1·5 (0–23) | 66 |
| Predominant channel substrate | cnm | Score totalb | RHQ | 33·5 (0–52) | 99 |
| Predominant flow | cft | Score totalb | RHQ | 32·5 (2–55) | 100 |
| Channel features | ct | Score totalb | RHQ | 1 (0–47) | 51 |
| Land use within 5 m | rl | Score totalb | RHQ | 46 (0–80) | 100 |
| Banktop vegetation structure | btv | Score totalb | RHQ | 21·5 (5·5–30) | 100 |
| Bankface vegetation structure | bfv | Score totalb | RHQ | 26·5 (4–30) | 100 |
| Channel vegetation types | cv | Score totalb | RHQ | 19 (0–43·6) | 97 |
| Land use within 50 m | lu | Score totalb | RHQ | 2·5 (0·6–8) | 100 |
| Natural bank profiles | bn | Score totalb | RHQ | 1·5 (0–3·5) | 91 |
| Extent of trees | rt | Score totalb | RHQ | 4·5 (0–5) | 99 |
| Shading of channel | rs | Score totalb | RHQ | 1 (0–2) | 94 |
| Overhanging boughs | rob | Score totalb | RHQ | 1 (0–2) | 89 |
| Exposed bankside roots | bbr | Score totalb | RHQ | 1 (0–2) | 74 |
| Underwater tree roots | bur | Score totalb | RHQ | 1 (0–2) | 56 |
| Fallen trees | bft | Score totalb | RHQ | 1 (0–2) | 74 |
| Coarse woody debris | cd | Score totalb | RHQ | 1 (0–2) | 83 |
| Flow types along 500 m | cf | Score totalb | RHQ | 5 (2–9) | 100 |
| Channel and bank features along 500 m | ff | Score totalb | RHQ | 3 (0–11) | 80 |
| Features of special interest along 500 m | fsi | Score totalb | RHQ | 1 (0–8) | 71 |
| Channel chocked with vegetationc | ccv | Score totalb | RHQ | 0 (0–1) | 2 |
| Predominant artificial bank material | bam | Score totalb | RHM | 2·5 (0–60) | 65 |
| Bank modifications | bm | Score totalb | RHM | 4·5 (0–70) | 73 |
| Artificial channel materialc | cam | Score totalb | RHM | 0 (0–10) | 7 |
| Channel modifications | cm | Score totalb | RHM | 0 (0–50) | 14 |
| Artificial bank profiles | ba | Score totalb | RHM | 2 (0–5) | 73 |
| Dam/weir | sd | Score totalb | RHM | 0 (0–5) | 28 |
| Bridge | sb | Score totalb | RHM | 0 (0–5) | 44 |
| Fordc | sf | Score totalb | RHM | 0 (0–2) | 8 |
| Deflectorc | sde | Score totalb | RHM | 0 (0–3) | 2 |
| Channel realignment | cmr | Score totalb | RHM | 0 (0–2) | 12 |
| Water impoundment by weir/dam | cmi | Score totalb | RHM | 0 (0–2) | 23 |
- Variable Groups: Typology, regional natural characteristics, RHQ, habitat quality features; RHM, habitat modification features.
- a For dummy variables, number of sites coded as ‘1’ is given.
- b Calculated score of individual features according to the SIHM method (Tavzes and Urbanič, 2009).
- c Variables excluded from further analysis owing to low occurrence frequency.

Data on morphological features (variables) were obtained along a 500-m-long stretch of the river using an adapted version of the UK RHS method (Raven et al., 2003, Tavzes and Urbanič, 2009). Bank and channel features (predominant substrate, physical features of channel and banks, flow type, channel vegetation type, land use, vegetation structure of banks and adjacent land) were recorded at 10 spot checks, spaced every 50 m. The sweep-up part of the survey along the whole stretch covers other features: land use in the 50-m stretch from the channel, bank profile, extent of trees, extent of bank and channel features, features of special interest and artificial features. The features recorded with the RHS method were upgraded in the SIHM method (Tavzes and Urbanič, 2009) where different weights were appointed to categories of each feature, depending on its influence on the benthic invertebrate community. Altogether, 33 features were recorded using the SIHM method (Table 1). RHQ features (RHQ variables) represent characteristics of habitat quality, including river channel and banks, riparian features and land use within 50 m from the channel. Higher values of RHQ variables indicate greater habitat diversity. RHM features (RHM variables) are derived from data on extent and impact of bank and channel modifications, which are weighted because of their impact on natural habitat. Higher values of RHM variables are an indication of bigger morphological alteration.
Data analysis
Preliminary morphological variables were examined with regard to occurrence frequency and gradient range. Variables present at less than 10% of sampling sites were excluded (Table 1). The remaining 29 morphological variables were used in the analyses – 21 and 8 from groups of RHQ features and RHM features, respectively. Regional natural characteristics were included in the analysis with eight variables. The region variables, karst spring influence and intermittency were coded as dummy (0/1) variables and river size as a variable with four classes. Altitude and slope were ln(x + 1)-transformed to approximate a normal distribution. River habitat variables were not transformed prior to analysis because the same unit score is used for all variables derived from the SIHM method (Tavzes and Urbanič, 2009). To investigate the relationship of environmental variables among and within analysed groups of variables, a Spearman rank correlation coefficient (RSp) was calculated for each pair of environmental variables using SPSS Statistics version 21.0 (IBM, 2012).
Ordination techniques were carried out to analyse associations among three groups of environmental variables, and between different groups of environmental variables and benthic invertebrate assemblages. Benthic invertebrate data were ln(x+1) transformed prior to analysis. All ordination techniques were conducted using the software package CANOCO 4.5 (ter Braak and Šmilauer, 2002); in all analyses, species were centred and standardized, and the option of downweighting rare species was enabled (emphasis was given to more commonly distributed species). When determining significant variables, the Monte Carlo permutation test (999 unrestricted permutations) with Bonferroni correction (α = 0·05/n, where n is the number of tests) was used. Firstly, a redundancy analysis with an automatic forward selection routine was used to investigate the significance and importance of typology variables in explaining variations of RHQ data or RHM data separately (Figure 2).
Furthermore, a detrended correspondence analysis performed on the benthic invertebrate dataset revealed the appropriateness of a unimodal response model (canonical correspondence analysis, CCA). The relationship between the environmental variables and benthic invertebrate data was therefore analysed using CCA (ter Braak and Prentice, 1988) and partial CCA (pCCA, Borcard et al., 1992). For the first overview of the environment–community relations, a CCA analysis with an automatic forward selection routine was applied on all environmental variables. This process tested the individual effects of each of the environmental variables (marginal effects) and the effect that each variable has, in addition to other selected variables (conditional effects) (Lepš and Šmilauer, 2003). Secondly, the importance of the three variable groups (typology, RHQ and RHM) in explaining variability among benthic invertebrate assemblages was tested using pCCA. pCCA allows partitioning of the variation in a species-sample data matrix, owing to the unique effects of explanatory variables and their combined effects. Prior to pCCA, variables were selected within each variable group with automatic forward selection in CCA analysis. The total explained variance among benthic invertebrate assemblages with forward-selected environmental variables from three groups was partitioned into (i) the variance uniquely explained by each variable group, (ii) the variance explained by combined effects of each pair of variable groups and (iii) the variance explained by combined effects of all three variable groups together.

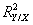 is the sample estimation of the assemblage variance
is the sample estimation of the assemblage variance 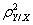 . The same number of samples and a similar number of predictors (between 5 and 12) in all the models lead to the very similar ratio of explained variance among the variable groups (regression curve; y = 0·76x − 0·0012, r2 = 0·977, P < 0·05).
. The same number of samples and a similar number of predictors (between 5 and 12) in all the models lead to the very similar ratio of explained variance among the variable groups (regression curve; y = 0·76x − 0·0012, r2 = 0·977, P < 0·05).Results
Relationships among environmental variables
Several statistically significant relationships (P < 0·05) between pairs of environmental variables were observed with Spearman rank correlation (RSp, Appendixes 1–6). Strong correlations (│RSp│ > 0·7) were observed only between variables within variable groups. In the RHQ group, land use within 5 m was strongly correlated with land use within 50 m and banktop vegetation structure; shading of channel also showed a strong correlation with overhanging boughs. The RHM group includes three variables among which a strong correlation was observed (predominant artificial bank material, bank modifications and channel modifications). All strong correlations were positive. Few variable pairs of different variable groups showed moderate correlations (0·50 < │RSp│ < 0·70). Slope was positively correlated with features of special interest along 500 m. Predominant natural bank material was negatively correlated with bank modifications. Other moderate correlations were observed within variable groups. In the typology group the Dinaric Western Balkan ecoregion was negatively correlated with the Alps ecoregion and the lowland region. Slope showed a positive correlation with the Alps ecoregion and a negative correlation with the river size class. In the RHQ group, some expected moderate correlations were observed. Predominant channel substrate correlated positively with predominant flow and predominant natural bank material. Also, among tree-associated variables (shading of channel, overhanging boughs, exposed bankside roots, fallen trees and coarse woody debris), positive moderate correlations were observed. Correlations between most pairs of variables were weak (│RSp│ < 0·5).
Associations among three groups of environmental variables
The associations among three groups of environmental variables were evaluated using the redundancy analysis. Typology variables explained 19% of the variability in RHQ variables (Figure 3a) with four statistically significant variables in the forward selection and only 7% of the variability in RHM variables (Figure 3b) with two statistically significant variables. In both datasets, statistically significant variables included slope and river size class. The most explanatory among typology variables was slope, explaining 9% and 3% of the variability in RHQ and RHM variables, respectively. Karst spring influence and intermittency were among the least explanatory variables in both datasets.

Benthic invertebrate responses to environmental variables
The total explained variance of 302 sites and 453 benthic invertebrate taxa dataset was 0·95 (27%). The highest explanatory power of individual variables was observed for slope (0·19), predominant flow and Alps ecoregion (0·17) (Table 2). Also, some other variables showed considerable explanatory power, 0·14–0·09 (predominant channel substrate, lowland region, river size class, features of special interest along 500 m and flow types along 500 m). The lowest explanatory power was observed for bridge (0·01). Among the RHM variables, the most explanatory was water impoundment by weir/dam (0·04). By using an automatic forward selection routine on all environmental variables, 16 statistically significant variables were selected (Table 2). The model of 16 selected variables explained 0·72 (21%) of the benthic invertebrate dataset (Figure 4). The 16 selected variables belong to all three groups, eight and seven variables were chosen from the RHQ and typology groups, respectively, and only one variable was chosen from RHM group. Of the 16 selected variables, only three variables from the RHQ group were not chosen when running the forward selection for each variable group separately (bank features, land use within 5 m, and banktop vegetation structure).
| Before FS | After FS all | After FS groups | ||||
|---|---|---|---|---|---|---|
| Environmental variable | Variable group | ƛ | ƛ | P | ƛ | P |
| Region: Lowland | Typology | 0·10 | 0·06 | 0·001 | 0·04 | 0·001 |
| Ecoregion: Alps | Typology | 0·17 | 0·05 | 0·001 | 0·08 | 0·001 |
| Ecoregion: Dinaric Western Balkan | Typology | 0·08 | ||||
| Catchment size class | Typology | 0·10 | 0·04 | 0·001 | 0·06 | 0·001 |
| Karst spring influence | Typology | 0·06 | 0·06 | 0·001 | 0·06 | 0·001 |
| Intermittency | Typology | 0·03 | 0·02 | 0·001 | 0·02 | 0·001 |
| Altitude | Typology | 0·08 | 0·02 | 0·001 | 0·03 | 0·001 |
| Slope | Typology | 0·19 | 0·19 | 0·001 | 0·19 | 0·001 |
| Predominant natural bank material | RHQ | 0·08 | 0·02 | 0·002 | ||
| Bank features | RHQ | 0·05 | 0·02 | 0·001 | ||
| Predominant channel substrate | RHQ | 0·14 | 0·03 | 0·001 | 0·04 | 0·001 |
| Predominant flow | RHQ | 0·17 | 0·11 | 0·001 | 0·17 | 0·001 |
| Channel features | RHQ | 0·02 | ||||
| Land use within 5 m | RHQ | 0·04 | 0·02 | 0·001 | ||
| Banktop vegetation structure | RHQ | 0·04 | 0·02 | 0·001 | ||
| Bankface vegetation structure | RHQ | 0·02 | 0·01 | 0·001 | 0·02 | 0·004 |
| Channel vegetation types | RHQ | 0·06 | 0·03 | 0·001 | 0·05 | 0·001 |
| Land use within 50 m | RHQ | 0·05 | 0·02 | 0·001 | ||
| Natural bank profiles | RHQ | 0·02 | ||||
| Extent of trees | RHQ | 0·04 | ||||
| Shading of channel | RHQ | 0·04 | 0·02 | 0·001 | 0·03 | 0·001 |
| Overhanging boughs | RHQ | 0·04 | ||||
| Exposed bankside roots | RHQ | 0·04 | 0·02 | 0·001 | ||
| Underwater tree roots | RHQ | 0·06 | 0·02 | 0·001 | ||
| Fallen trees | RHQ | 0·04 | ||||
| Coarse woody debris | RHQ | 0·03 | ||||
| Flow types along 500 m | RHQ | 0·09 | 0·02 | 0·003 | ||
| Channel and bank features along 500 m | RHQ | 0·08 | 0·02 | 0·001 | ||
| Features of special interest along 500 m | RHQ | 0·09 | 0·06 | 0·001 | ||
| Predominant artificial bank material | RHM | 0·02 | 0·03 | 0·001 | ||
| Bank modifications | RHM | 0·03 | 0·03 | 0·001 | ||
| Channel modifications | RHM | 0·02 | ||||
| Artificial bank profiles | RHM | 0·03 | 0·02 | 0·003 | ||
| Dam/weir | RHM | 0·02 | 0·03 | 0·001 | ||
| Bridge | RHM | 0·01 | ||||
| Channel realignment | RHM | 0·02 | ||||
| Water impoundment by weir/dam | RHM | 0·04 | 0·02 | 0·002 | 0·04 | 0·001 |
- Independent effects: before FS, before forward selection. Conditional effects: after FS all, after forward selection with all variables; after FS groups, after forward selection within each variable group. For variable groups (Typology, RHQ, RHM) see Table 1.

Variance partitioning of three groups of environmental variables and benthic invertebrate assemblages
Variance partitioning was run with 24 variables selected after forward selection routine for each variable group separately. Three variable groups consisted of 5–12 selected variables (Table 2), with the highest number in the RHQ group (12 out of 21), followed by the typology group (seven out of eight) and RHM group (five out of eight). All 24 selected variables together explained 24% of the benthic invertebrate assemblages' variability. Clearly, unique effects of variable groups were more important in explaining variability in the benthic invertebrate composition than joint effects (69% and 31% of the explained variance, respectively, Figure 5). A similar share of unique effects was explained by the typology (30%) and RHQ (31%) groups, but the explanatory power of the RHM group was considerably smaller (8%). Among joint effects, the interaction between the typology and RHQ groups was most important, accounting for 20% of explained variance. Other interactions between pairs of groups were less important. Joint effects of all three variable groups explained 6% of variability.

Discussion
The relation of regional natural characteristics to local morphological features
The occurrence and shape of local river morphological features depend on regional factors (Frissell et al., 1986; Poff, 1997). Regional natural characteristics (typological variables) in Slovenian rivers explained a considerable amount of variation in local habitat quality features (RHQ). The highest explanatory power among typological variables was observed for slope, followed by river size class and Alps ecoregion. Slope is commonly recognized as an important parameter structuring channel morphology (Allan, 2004), and the thesis is confirmed also by low, but mostly significant, positive correlations between slope and most of the RHQ variables in our study. Steeper slopes are mostly found in upland regions and connected to narrow valleys with small rivers. Tavzes and Urbanič (2009) reported that Slovenian rivers of the Alps ecoregion have habitat quality characteristics typical of alpine streams – related to high-energy flows (Szoszkiewicz et al., 2006). Hence, the significance of the Alps ecoregion in our analysis was not surprising. The significance of river size partly depends on highly energetic streams of the Alps ecoregion, influencing river flow and channel substrate heterogeneity, but the negative correlation of river size to riparian zone quality features also suggests more naturalness with smaller river size.
Because a relationship between regional factors and local RHQ characteristics exists, regional factors could define the alteration of river morphological habitat to some extent. Alpine regions with narrow and remote river valleys are less suitable for human settlement. However, these attributes are becoming more and more interesting for different human activities (e.g. tourism and hydropower plants), and high-energy flows of alpine rivers causing in-stream instability represent an obstacle. For this reason, many streams in the Alpine area are now affected by morphological alterations, such as resectioning and reinforcement of banks, bridges and weirs (Bona et al., 2008; Tavzes and Urbanič, 2009; Wyżga et al., 2011). Lowland rivers, on the other hand, have wider riparian zones, mostly connected to large floodplains. Human occupation of floodplains and riparian zones lead to a wide range of morphological alteration affecting lowland rivers (Pedersen, 2009; Pedersen and Friberg, 2009), such as straightening and deepening. Therefore, morphological degradation of river habitats occurs irrespective of regional factors, just as its ground reason differs. This thesis was confirmed in our study, as the typological variables explained an almost negligible amount of variation in local habitat modification features. A significant, although low, explanatory power was observed for slope and river size class. Whereas slope represents mostly a regional factor, by being steeper in upland and gentler in lowland regions, river size in Slovenia is not merely a regional factor, as small streams with their sources up to large rivers are found in mountainous and lowland regions. However, river size is still partly related to bank modifications. One possible reason for this correlation is the social view of larger rivers as more threatening, causing more erosion or flooding and thus requiring more regulation structures. The significance of river size class in the present study also demonstrates the nonregional presence of morphological modification.
Linking morphological features to the response of benthic invertebrate assemblages
The SIHM method was developed for the assessment of river habitat quality (RHQ) and modification (RHM) features. Higher values of RHQ features indicate a high diversity of habitat, whereas severe morphological degradation is characterized by high values of RHM features. We tested individual features of the SIHM method for their significance in structuring benthic invertebrate assemblages. Generally, habitat quality features revealed more explanatory power than habitat modification features. Among RHQ features, the highest explanatory power was observed for predominant flow and predominant channel substrate – the two variables most commonly observed in affecting aquatic assemblages (Richards et al., 1993; Lammert and Allan, 1999; Sandin, 2003; Sandin and Johnson, 2004, Syrovátka et al., 2009). The strong and more diversified river flow mostly relates to coarse and heterogeneous channel substrate (Statzner and Higler, 1986; Poff et al., 1997), but these variables were only moderately correlated in our study. Moreover, predominant flow compared with predominant channel substrate showed higher explanatory power in structuring benthic assemblages. River flow represents a direct physical force affecting organisms, but its indirect effects are also significant for benthic assemblages, including oxygen content of the stream, food delivery and substratum composition. Studies comparing the effects of both flow and substrate variables are rare (Urbanič et al., 2005; Friberg et al., 2009a; Sandin, 2009; Wyżga et al., 2011), but all revealed partly different relations of predominant flow and substrate to benthic assemblages. Besides flow, other mechanisms affect substrate composition and consequently biota (Wyżga et al., 2011). Different in-stream structures interrupting longitudinal continuity upstream can reduce the availability of substrate for fluvial transport (Kondolf, 1997), and the local riparian management can affect in-stream substrate and biota by the reduction or acceleration of bank erosion and by restraining the sediment input from the catchment (Allan, 2004). In fact, our study showed a positive correlation of channel substrate to the structure of riparian vegetation and land use.
Riparian vegetation is another habitat feature commonly exerting great influence upon habitat diversity and, consequently, the structure and function of aquatic assemblages (Cummins et al., 1989; Bis et al., 2000; Sandin, 2009). The SIHM method comprises several features describing riparian vegetation structure, and although all features significantly contribute to structuring benthic invertebrate assemblages, the highest explanatory power observed was still threefold lower than that of predominant flow and channel substrate. Also, in-stream vegetation that plays a vital role in increasing habitat diversity and creating aquatic refugia for biota (Sandin and Johnson, 2004; Pinto et al., 2006) explained as low amount of benthic assemblages' variability as riparian vegetation features. Our results suggest that the structure of benthic assemblages depends more on flow and substrate characteristics than on riparian and in-stream vegetation structure. These findings are important guidance for river management, as riparian vegetation reestablishment is one of the most common river rehabilitation practices today (Death and Collier, 2009; Riley and Dodds, 2012). Also, Greenwood et al. (2012) imply that poor in-stream habitat, due to changed flow dynamics and sedimentation, can limit the effectiveness of riparian management. On the other hand, Urban et al. (2006) found that riparian vegetation was a stronger predictor of benthic invertebrate community than habitat and reach in-stream variables, the latter including substrate and stream discharge. However, the habitat, reach and riparian spatial scale were much smaller than in the present study (up to 150 m). The importance of flow and substrate diversity in our study was also reflected in reach-scale variables (500 m), with the explanatory power higher than that of any riparian vegetation feature.
Human modifications of river habitat are reflected in RHM features, all of them showing very low explained variability of benthic invertebrate assemblages. The possible reason for a lower explanatory power of RHM than of RHQ features is the nature of the survey, where RHM features might be as equally present as absent, in contrast to RHQ features that can almost always be assigned to one of the categories (Raven et al., 1997; Tavzes and Urbanič, 2009). However, even frequent RHM features did not explain a greater amount of assemblages' variability across the whole study area. The most important RHM feature seems to be water impoundment by dam/weir, in agreement with the findings of Marzin et al. (2012), where the presence of an impoundment emerged as the main human pressure factor shaping the fish and macroinvertebrate assemblages at the reach scale. Similar to the most explanatory RHQ variable (predominant flow), the change of flow characteristics caused by impoundment affects other morphological features (e.g. channel substrate) and results in direct and indirect effects on benthic invertebrate assemblages. On the other hand, studies across Europe identified bank resectioning and reinforcement as the most typical modification of rivers (Feld, 2004; Szoszkiewicz et al., 2006). The correlation of bank modification to benthic invertebrate metrics, observed by Erba et al. (2006), and the feature being one of the best explanatory RHM variables in our study confirm the importance of bank beside channel alteration.
The low explanatory power of morphological modification features might also reflect the overriding effect of habitat quality features. To some extent, RHQ features exert human influence. Namely, bank modification, such as straightening, influences predominant flow and channel substrate, as well as channel and bank natural features (Negishi et al., 2002; Pedersen, 2009). Also, riparian zone clearing lowers the values of riparian vegetation features and, owing to lessened shadowing, accelerates growth of in-stream vegetation (Julian et al., 2011). Additionally, a four times higher explanatory power of predominant flow, in comparison with that of water impoundment by dam/weir, was observed. According to the results of the present study, a higher importance in structuring benthic assemblages is suggested for the habitat quality and its diminishment by human influence than for morphological alteration features itself.
Co-influence of regional natural characteristics and local morphological features
As observed before, habitat features on local scales might be influenced by regional parameters (typology), and hence, the real effect on aquatic assemblages might be obscured (Richards et al., 1997; Cortes et al., 2009). The issue of hierarchical interaction of multiscale factors has been of special concern for studies relating natural characteristics and human alterations to river communities, with authors emphasizing the importance of larger scales (Roth et al., 1996; Lammert and Allan, 1999) or of smaller scales (Ormerod et al., 1993; Richards et al., 1993; Heino et al., 2004). Marginal effects of different scale variables in the present study revealed a similar explanatory power of the best local habitat quality features (predominant flow and channel substrate) and best regional natural variables (Alps ecoregion and slope). Moreover, using variance partitioning, we observed almost even distinct effects of typological and local habitat quality variables. This implies the importance of quality parameters at both scales, which is in accordance with the findings of other authors (Brosse et al., 2003; Sandin, 2003; Johnson et al., 2007). The observed amount of joint effects between habitat quality features and typological parameters was considerable but lower than that of distinct effects. The interaction of parameters on both scales, and aforementioned explanatory power of regional natural parameters on habitat quality characteristics, lead to two definitive conclusions – the larger-scale parameters constrain the processes on smaller scales to a certain extent, and the resulting physical patterns influence the biology of stream, which is agreed also by other authors (Richards et al., 1997; Verdonschot, 2006; Li et al., 2012, Marzin et al., 2012). But the observed large share of distinct effects implies that there is a considerable part of benthic invertebrate assemblage variability in Slovenia dependent on habitat quality features irrespective of typological characteristics.
In contrast to habitat quality features, we cannot make any presumptions for habitat modification features, as either their distinct or joint effects with typological parameters were too low. One of the possible reasons is, on the one hand, the high diversity of regional and local characteristics of Slovenian landscape, influencing the adjustment of benthic assemblages and, on the other hand, the presence of a wide variety of modification features irrespective of the region. As benthic assemblages are adapted to the former natural conditions, similar modification features might result in different effects with regard to regional characteristics. Bank stabilization by concrete or rip-rap in alpine rivers, with natural banks consisting of rocks or stones, results in effects different than those in lowland rivers, where naturally, sand or earth forms bank profiles. Consequently, using the dataset from different regions jointly might obscure the effects of morphological modification.
Conclusions
Several conclusions can be drawn from our analysis of RHQ and RHM features, according to the Slovenian hydromorphological (SIHM) assessment method. In our study, a large heterogeneous area was sampled. Therefore, we did not expect the great explanatory power of local morphological features but aimed at defining general patterns. The most important local habitat characteristics for structuring benthic invertebrate assemblages were identified, with predominant flow and channel substrate as the best explanatory variables. Almost all habitat quality features proved important, but the effects of habitat modification features were marginal. We suggest that benthic invertebrate assemblages respond less to the physical alteration itself and more to the effect that the alteration exerts on habitat quality features. Moreover, local habitat quality features depend on regional characteristics, but only to some extent. When discussing management options, general guidelines are most welcome, but even more desirable are their high confidence levels. To lessen the effect of regional parameters, analyses on regional scale should be conducted. Because our study revealed the great importance of ecoregions for structuring benthic assemblages, we propose further analyses on the ecoregion scale. With the whole gradient of habitat alteration covered, we also expect these analyses within more homogeneous natural habitat to more clearly reveal the significance of modification features. Finally, the most relevant morphological features, according to our findings, are directly connected to river flow. Hence, for more effective planning of management practices, we suggest further study for more detailed determination of the interaction between hydrological and morphological parameters on aquatic assemblages.
Acknowledgements
This study was supported by the Ministry of Agriculture and the Environment of the Republic of Slovenia as a part of the national programme for the implementation of the EU Water Framework Directive. We would also like to thank the anonymous reviewer and the editor for their useful comments to a previous version of the manuscript.
Appendix 1: Statistically significant Spearman's correlation coefficients (RSp) for the combinations of environmental variables between the typology and RHQ groups.
| Variable | ER3.11 | ER4 | ER5 | Size_cl | Kspring | Intermit | Alt | Slope |
|---|---|---|---|---|---|---|---|---|
| bnm | −0·272** | 0·343** | −0·138* | 0·150** | 0·119* | 0·476** | ||
| bf | −0·176** | 0·191** | −0·162** | 0·359** | ||||
| cnm | −0·135* | 0·376** | −0·230** | 0·141* | 0·165** | 0·426** | ||
| cft | −0·120* | 0·472** | −0·333** | −0·261** | 0·266** | 0·394** | ||
| ct | −0·158** | 0·163** | 0·292** | |||||
| rl | −0·131* | 0·236** | −0·156** | 0·144* | 0·242** | |||
| btv | 0·161** | 0·121* | 0·172** | |||||
| bfv | −0·113* | 0·167** | ||||||
| cv | −0·207** | 0·282** | 0·131* | 0·308** | −0·162** | |||
| lu | −0·139* | 0·198** | −0·127* | 0·142* | 0·179** | 0·236** | ||
| bn | 0·138* | |||||||
| rt | −0·145* | 0·222** | 0·136* | 0·250** | ||||
| rs | 0·167** | −0·464** | 0·332** | |||||
| rob | 0·184** | −0·401** | 0·283** | |||||
| bbr | −0·241** | 0·248** | −0·241** | |||||
| bur | −0·366** | 0·322** | −0·203** | −0·187** | ||||
| bft | −0·159** | 0·202** | −0·278** | 0·213** | ||||
| cd | −0·238** | 0·229** | ||||||
| cf | −0·255** | 0·279** | −0·244** | 0·219** | 0·445** | |||
| ff | −0·237** | 0·203** | −0·241** | 0·127* | 0·471** | |||
| fsi | −0·311** | 0·308** | −0·468** | 0·317** | 0·597** |
- RSp > 0·5 are in bold.
- * P < 0·05,
- ** P < 0·01.
Appendix 2: Statistically significant Spearman's correlation coefficients (RSp) for the combinations of environmental variables between the typology and RHM groups.
| Variable | ER3.11 | ER4 | ER5 | Size_cl | Kspring | Intermit | Alt | Slope |
|---|---|---|---|---|---|---|---|---|
| bam | 0·124* | −0·124* | 0·238** | −0·181** | −0·138* | |||
| bm | 0·194** | 0·224** | −0·170** | −0·247** | ||||
| cm | −0·128* | 0·162** | 0·122* | −0·134* | ||||
| ba | 0·169** | −0·162** | 0·240** | −0·174** | −0·167** | |||
| sd | −0·181** | −0·163** | 0·119* | −0·142* | 0·137* | |||
| sb | ||||||||
| cmr | ||||||||
| cmi | 0·134* | 0·135* | −0·172** |
- * P < 0·05,
- ** P < 0·01.
Appendix 3: Statistically significant Spearman's correlation coefficients (RSp) for the combinations of environmental variables between the RHM and RHQ groups.
| Variable | bam | bm | cm | ba | sd | sb | cmr | cmi |
|---|---|---|---|---|---|---|---|---|
| bnm | −0·494** | −0·563** | −0·211** | −0·442** | −0·189** | −0·268** | −0·329** | −0·131* |
| bf | −0·202** | −0·316** | −0·124* | −0·219** | −0·136* | −0·183** | −0·182** | |
| cnm | −0·191** | −0·148* | −0·223** | −0·205** | ||||
| cft | 0·169** | 0·164** | −0·302** | |||||
| ct | −0·199** | −0·138* | −0·197** | |||||
| rl | −0·420** | −0·458** | −0·189** | −0·362** | −0·219** | −0·319** | −0·253** | −0·154** |
| btv | −0·423** | −0·433** | −0·188** | −0·346** | −0·243** | −0·298** | −0·322** | −0·175** |
| bfv | −0·400** | −0·422** | −0·219** | −0·351** | −0·262** | −0·319** | −0·294** | −0·148** |
| cv | ||||||||
| lu | −0·324** | −0·390** | −0·222** | −0·271** | −0·233** | −0·394** | −0·247** | −0·164** |
| bn | −0·139* | −0·246** | −0·162** | −0·154** | −0·209** | −0·232** | ||
| rt | −0·301** | −0·360** | −0·206** | −0·269** | −0·136* | −0·250** | −0·269** | |
| rs | −0·215** | −0·204** | −0·214** | |||||
| rob | −0·257** | −0·260** | −0·250** | −0·122* | ||||
| bbr | −0·356** | −0·341** | −0·153** | −0·334** | −0·134* | −0·145* | ||
| bur | −0·205** | −0·139* | −0·143* | |||||
| bft | −0·405** | −0·421** | −0·361** | −0·182** | ||||
| cd | −0·292** | −0·319** | −0·124* | −0·249** | −0·133* | |||
| cf | −0·154** | −0·125* | −0·158** | |||||
| ff | −0·262** | −0·376** | −0·144* | −0·292** | −0·127* | −0·116* | −0·208** | −0·187** |
| fsi | −0·359** | −0·410** | −0·200** | −0·338** | −0·151** | −0·193** | −0·319** |
- RSp > 0·5 are in bold.
- * P < 0·05,
- ** P < 0·01.
Appendix 4: Statistically significant Spearman's correlation coefficients (RSp) for the combinations of environmental variables within the typology group.
| Variable | ER3.11 | ER4 | ER5 | Size_cl | Kspring | Intermit | Alt |
|---|---|---|---|---|---|---|---|
| ER4 | −0·400** | ||||||
| ER5 | −0·518** | −0·576** | |||||
| Size_cl | 0·358** | −0·177** | −0·154** | ||||
| Kspring | −0·299** | 0·224** | |||||
| Intermit | −0·137* | −0·153** | 0·265** | −0·141* | |||
| Alt | −0·435** | 0·492** | −0·340** | 0·154** | |||
| Slope | −0·390** | 0·502** | −0·120* | −0·618** | 0·362** |
- RSp > 0·5 are in bold.
- * P < 0·05,
- ** P < 0·01.
Appendix 5: Statistically significant Spearman's correlation coefficients (RSp) for the combinations of environmental variables within the RHM group.
Appendix 6: Statistically significant Spearman's correlation coefficients (RSp) for the combinations of environmental variables within the RHM group.
| Variable | bnm | bf | cnm | cft | ct | rl | btv | bfv | cv | lu | bn | rt | rs | rob | bbr | bur | bft | cd | cf | ff |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| bf | 0·409** | |||||||||||||||||||
| cnm | 0·536** | 0·325** | ||||||||||||||||||
| cft | 0·252** | 0·255** | 0·563** | |||||||||||||||||
| ct | 0·419** | 0·360** | 0·398** | 0·291** | ||||||||||||||||
| rl | 0·482** | 0·261** | 0·310** | 0·263** | ||||||||||||||||
| btv | 0·395** | 0·227** | 0·267** | 0·212** | 0·763** | |||||||||||||||
| bfv | 0·363** | 0·166** | 0·175** | 0·141* | 0·414** | 0·594** | ||||||||||||||
| cv | −0·193** | −0·118* | −0·206** | −0·193** | ||||||||||||||||
| lu | 0·464** | 0·250** | 0·341** | 0·122* | 0·240** | 0·762** | 0·645** | 0·371** | −0·144* | |||||||||||
| bn | 0·298** | 0·136* | 0·115* | 0·143* | 0·129* | 0·161** | 0·115* | |||||||||||||
| rt | 0·429** | 0·289** | 0·243** | 0·199** | 0·217** | 0·494** | 0·556** | 0·644** | −0·247** | 0·480** | 0·165** | |||||||||
| rs | 0·123* | −0·115* | 0·188** | |||||||||||||||||
| rob | 0·124* | −0·142* | 0·157** | 0·304** | 0·799** | |||||||||||||||
| bbr | 0·141* | 0·172** | −0·203** | −0·146* | 0·144* | 0·122* | 0·193** | 0·586** | 0·628** | |||||||||||
| bur | −0·251** | −0·230** | −0·146* | 0·424** | 0·483** | 0·602** | ||||||||||||||
| bft | 0·283** | 0·242** | 0·173** | 0·273** | 0·151** | 0·169** | 0·193** | 0·145* | 0·273** | 0·536** | 0·546** | 0·574** | 0·429** | |||||||
| cd | 0·236** | 0·278** | 0·134* | 0·185** | 0·185** | 0·270** | 0·488** | 0·508** | 0·572** | 0·356** | 0·659** | |||||||||
| cf | 0·344** | 0·446** | 0·446** | 0·380** | 0·518** | 0·279** | 0·210** | −0·137* | 0·243** | 0·285** | 0·190** | 0·146* | 0·242** | 0·264** | ||||||
| ff | 0·539** | 0·728** | 0·378** | 0·280** | 0·597** | 0·352** | 0·223** | 0·147* | −0·149** | 0·311** | 0·328** | 0·278** | 0·229** | 0·296** | 0·378** | 0·319** | 0·617** | |||
| fsi | 0·487** | 0·467** | 0·359** | 0·373** | 0·399** | 0·405** | 0·321** | 0·188** | −0·131* | 0·333** | 0·390** | 0·362** | 0·330** | 0·221** | 0·318** | 0·315** | 0·603** | 0·651** |
- RSp > 0·5 are in bold.
- * P < 0·05,
- ** P < 0·01.

