

Stable coexistence of incompatible Wolbachia along a narrow contact zone in mosquito field populations
Abstract
In arthropods, the intracellular bacteria Wolbachia often induce cytoplasmic incompatibility (CI) between sperm and egg, which causes conditional embryonic death and promotes the spatial spread of Wolbachia infections into host populations. The ability of Wolbachia to spread in natural populations through CI has attracted attention for using these bacteria in vector-borne disease control. The dynamics of incompatible Wolbachia infections have been deeply investigated theoretically, whereas in natural populations, there are only few examples described, especially among incompatible infected hosts. Here, we have surveyed the distribution of two molecular Wolbachia strains (wPip11 and wPip31) infecting the mosquito Culex pipiens in Tunisia. We delineated a clear spatial structure of both infections, with a sharp contact zone separating their distribution areas. Crossing experiments with isofemale lines from different localities showed three crossing types: wPip11-infected males always sterilize wPip31-infected females; however, while most wPip31-infected males were compatible with wPip11-infected females, a few completely sterilize them. The wPip11 strain was thus expected to spread, but temporal dynamics over 7 years of monitoring shows the stability of the contact zone. We examined which factors may contribute to the observed stability, both theoretically and empirically. Population cage experiments, field samples and modelling did not support significant impacts of local adaptation or assortative mating on the stability of wPip infection structure. By contrast, low dispersal probability and metapopulation dynamics in the host Cx. pipiens probably play major roles. This study highlights the need of understanding CI dynamics in natural populations to design effective and sustainable Wolbachia-based control strategies.
Introduction
Wolbachia are maternally inherited intracellular α-proteobacteria that infect many arthropod species (Werren et al. 2008). Wolbachia are the most frequent bacterial endosymbiont described so far in insects, infecting 20–70% of species (Werren et al. 1995; Jeyaprakash & Hoy 2000; Hilgenboecker et al. 2008; Zug & Hammerstein 2012). This evolutionary success is mainly attributed to the ability of Wolbachia to manipulate the host reproductive system to its own advantage (Werren et al. 2008). The most common manipulation is cytoplasmic incompatibility (CI), which triggers embryonic mortality when infected males mate with uninfected females, whereas the reciprocal cross remains compatible. Thus, infected females have a reproductive advantage in polymorphic populations which allows the spread of infections. Cases of such spreads were reported in field populations of Drosophila simulans in California (Turelli & Hoffmann 1991), planthopper Laodelphax striatellus (Hoshizaki & Shimada 1995) and butterfly Eurema hecabe in Japan (Hiroki et al. 2005).
Wolbachia has attracted much attention as a promising tool to control diseases transmitted by mosquitoes, after the observation that some infections of Aedes or Anopheles mosquitoes by Wolbachia reduce the development time of their host and reduce the transmission of human pathogens such as the dengue and chikungunya viruses (Moreira et al. 2009; Walker et al. 2011; Blagrove et al. 2012) or Plasmodium (Kambris et al. 2010; Bian et al. 2013). Furthermore, Wolbachia-infected Aedes aegypti released in Australia have readily spread in field populations through CI, as predicted by infection dynamic models (Hoffmann et al. 2011). Better knowledge of CI dynamics is thus required to explore the full potential of Wolbachia in vector control strategies.
Wolbachia dynamics is made more complex by the presence of several incompatible strains. When males and females are infected with different and incompatible strains, CI occurs and can follow either unidirectional or bidirectional patterns (Riegler & Stauffer 2002; Mercot & Charlat 2004; Duron et al. 2006a; Atyame et al. 2014). Wolbachia dynamics has been largely explored theoretically, and three key models parameters describe how CI could influence the spread of Wolbachia in a panmictic host population (see Engelstadter & Telschow 2009 for review): (i) the transmission rate—that is the proportion of infected offspring produced by an infected female; (ii) the CI mortality (or CI level)—that is the proportion of offspring that die in incompatible crosses; and (iii) the effect of Wolbachia infection on female fecundity—that is on its host fitness (Fine 1978; Hoffmann et al. 1990; Turelli & Hoffmann 1995).
Incompatible Wolbachia strains cannot stably coexist in an unstructured and panmictic host population, as only compatible strains can resist the CI-driven competition (Rousset et al. 1991; Engelstadter & Telschow 2009). The pattern of CI will determine the competition outcome: in the case of bidirectional CI, the most frequent Wolbachia strain always outcompetes the rarest, while with unidirectional CI, the CI-inducing strain is expected to invade the population, once above an initial frequency threshold (Caspari & Watson 1959; Engelstadter & Telschow 2009). Only two studies report on Wolbachia strains competing in naturally infected host populations: (i) the cherry fruit fly Rhagoletis cerasi, in which flies coinfected with two Wolbachia strains invaded populations carrying only one Wolbachia infection in southern Europe (Riegler & Stauffer 2002), and (ii) D. simulans, in which the Wolbachia strain wRi invaded populations infected with the strain wAu on the east coast of Australia (Kriesner et al. 2013). In this latter case, data were best explained by postulating that both wAu and wRi increased females fecundity, highlighting a potential role for selection in the global distribution of Wolbachia.
However, invasion thresholds deduced for isolated panmictic populations cannot be directly applied to structured host populations connected by dispersal, as system dynamics and equilibrium states are modified (Barton & Turelli 2011; Hancock & Godfray 2012). Incompatible Wolbachia strains can indeed stably coexist in structured parapatric host populations if dispersal is below a critical threshold (Telschow et al. 2005; Flor et al. 2007).
Culex pipiens complex mosquitoes—whose main subspecies are Cx. p. quinquefasciatus and Cx. p. pipiens, ubiquitous in tropical and temperate regions, respectively—present an extreme situation. In temperate regions, two Cx. p. pipiens forms coexist (the pipiens and molestus forms), showing behavioural, physiological and genetic specificities (Fonseca et al. 2004). Culex pipiens quinquefasciatus and Cx. p. pipiens mosquitoes are naturally infected by Wolbachia strains (wPip) that belong to a unique clade of the B supergroup (Rasgon & Scott 2003; Duron et al. 2005, 2006b). However, wPip strains display a high genetic polymorphism at a small evolutionary scale (Duron et al. 2006b; Atyame et al. 2011a; Dumas et al. 2013) and complex CI patterns (Laven 1967a; Duron et al. 2006a; Atyame et al. 2011b, 2014). A recent survey showed that considerable genetic diversity is stably maintained within geographically close Cx. pipiens populations infected by compatible wPip strains (Duron et al. 2011).
We report here on a field situation in North Africa where Cx. pipiens populations were found infected by only two molecular strains, wPip31 in Algeria and wPip11 in Tunisia (Duron et al. 2006b). These two strains were defined using a set of polymorphic molecular markers and laboratory crosses showed CI between them (Duron et al. 2006a). This situation was thus a very attractive opportunity to study their evolutionary dynamics in natural populations. We have now mapped the distribution areas of these wPip strains in Algeria and Tunisia, characterized their CI patterns from isofemale lines sampled in different localities and analysed CI expression from direct observations in the field. We have still detected only the same two molecular wPip strains. However, we have found three strains in term of CI properties. wPip11-infected males always sterilize wPip31-infected females. While most wPip31-infected males were compatible with wPip11-infected females, a few completely sterilized them. In these conditions, the wPip11 strain is expected to outcompete the wPip31 strains over time. We have thus performed a 7-year follow-up of the wPip infections in field populations and have found that the contact zone between wPip11 and wPip31 strains has remained stable. By developing a spatially explicit model that better describes the field situation, we have identified critical parameters that might explain the long-term coexistence of these incompatible wPip infections.
Materials and methods
Mosquito collection and lines maintenance
Culex pipiens mosquitoes were collected as larvae or pupae in ten localities of Algeria (in 1997 and in June 2006 and 2008) and in 63 localities of Tunisia (in June 1996, 1997, 2003, 2005, 2008, 2009 and 2010, October 2010 and June 2011). All localities were not sampled each year (Table S1, Supporting information). Each sample was reared to adulthood in the laboratory, and approximately 100 specimens were stored at −20 °C or in liquid nitrogen until Wolbachia genotyping. Adults from 19 localities were allowed to mate to establish isofemale lines (Table S1, Supporting information). Each egg raft (containing 100–300 eggs) was individually isolated for hatching, and the genotyping of its Wolbachia infection was performed by analysing two first-instar larvae (L1). All isofemale lines (n = 245) were reared in 65-dm3 screened cages kept in a single room at 22–25 °C, under a 12-h light/12-h dark cycle. Larvae were fed with a mixture of shrimp powder and rabbit pellets, while adults were fed with honey solution.
Molecular typing
Wolbachia genotyping was performed using two wPip polymorphic markers (among several previously described to differentiate the wPip strains): the ankyrin domains gene ank2 (Duron et al. 2007) and the DNA mismatch repair protein gene MutL (Atyame et al. 2011a). These two genes differentiate the wPip strains investigated here on the basis of the size of the PCR-amplified fragments: ank2 displays 313- and 511-bp fragment sizes, and MutL displays 374- and 437-bp fragment sizes for the strains wPip11 and wPip31, respectively (Fig. S1, Supporting information).
We also analysed mitochondrial variability by sequencing 852 bp of the cytochrome b gene (cytb) in 12 isofemale lines from Algeria and Tunisia: five wPip11 lines from four localities (Sousse, #25; Zerga, #30; Sokra, #37 and Aïn Tounga, #51) and seven wPip31 lines from seven localities (Harash, #3; Guelma, #6; Douas, #8; Kala, #9; Tabarka, #31; Kef1, #34; Boussalem1, #42).
Finally, we checked for a relationship between Cx. pipiens forms (form pipiens and form molestus) and wPip infection. The CQ11 microsatellite locus (Bahnck & Fonseca 2006) was used to distinguish the two forms in samples from two wPip11 localities (Ayed, #24 and Riadh, #27) and two wPip31 localities (Tabarka, #31 and Boussalem2, #71). The CQ11 microsatellite loci were previously found effective for molecular identification of pipiens and molestus forms, as well as hybrids, in Cx. pipiens mosquitoes in Morocco (Amraoui et al. 2012).
DNA was extracted from adult mosquitoes and larvae using a cetyltrimethylammonium bromide (CTAB) protocol (Rogers & Bendich 1988). All PCRs were performed with 50 ng of genomic DNA solution in a 40 μL final under the following conditions: 94 °C for 30 s, 52 °C for 30 s and 72 °C for 1–1.5 min for a total of 33 cycles (primers are listed in Table S2, Supporting information). Amplified DNA fragments were separated by agarose gel (1.5%) electrophoresis. For sequencing, PCR products were purified with the QIAquick gel extraction kit (QIAGEN, Valencia, CA, USA) and then directly sequenced with an ABI Prism 3130 sequencer using the BigDye Terminator kit (Applied Biosystems).
Cytoplasmic incompatibility
In the laboratory
Crosses were carried out with 25–50 virgin females and an equivalent number of males derived from isofemale lines. A first set of crosses was performed between wPip11 and wPip31 lines isolated from localities where only one Wolbachia molecular strain was present: five wPip11 lines (three localities: Tunis, #16; Sousse, #25; and Sokra, #37) and 16 wPip31 lines (eight localities: Tabarka, #31; Kef1, #34; and Boussalem1, #42 from Tunisia and Guelma, #6; Lac, #7; Douas, #8; Kala, #9; and Souk Ahras, #10 from Algeria). Next, crosses were performed between wPip11 and wPip31 lines from the same locality, that is between two wPip11 and two wPip31 isofemale lines isolated from the Tunisian samples Zerga (#30) and Ain Tounga (#51). To assess CI phenotypes among wPip11-infected males, isofemale-derived wPip11 males from three localities (El Battan, #43; El Manar, #46; and Aïn Tounga, #51) were crossed with wPip31 females derived from the Har isofemale line (Harash, #3). All individuals were 2–5 days old. Females were allowed to blood-feed 5 days after caging, and their egg rafts were collected after 5 days and stored individually until hatching. The CI status of each cross was determined by examining egg hatching rate (HR) under a binocular microscope. All unhatched egg rafts were checked for fertilization through observation of embryonic development as described by Duron & Weill (2006).
Mating preferences of wPip11 and wPip31 mosquitoes were measured in laboratory cages (65 dm3), where 100 males and 100 females from a wPip11 isofemale line were placed with an equivalent number of males and females from a wPip31 isofemale line (later identified as a wPip31_U line). Two types of confrontations were set up: Sok × Kef1-1 and Sok × Kef1-2, both showing unidirectional CI (Sok males sterilizing kef1-1 and kef1-2 females). Three replicates were performed for each type of confrontation, so a total of six cages were set up. All individuals were 1 day old and virgin, and all the mosquitoes were introduced into the cage at the same time. Females were blood-fed 5 days after caging and allowed to oviposit on a water cup. Egg rafts were individually stored and were analysed as described above.
In field populations
The expression of CI in Cx. pipiens natural populations was assessed by sampling egg rafts in Tunisian localities in 2009 and 2010. The egg rafts were carefully removed with a paintbrush from the surface of stagnant water, placed separately in 24-well plates and brought to the laboratory to observe hatching. As larvae hatch within 36–48 h after ovoposition at 25 °C, HR was evaluated >72 h after collection under a binocular microscope and egg rafts that had already hatched in the field were easy to qualify.
Population cages
The invasion dynamics of the wPip11 strain was examined in population cages (65 dm3) in laboratory through confrontations between the wPip11 isofemale line Tn and the wPip31 isofemale line Har (a wPip31_U line), showing unidirectional CI (Tn males sterilizing Har females). To avoid side effects of nuclear genomes, the cytoplasms (including wPip strains) of the Tn and Har lines were introduced into the same nuclear background (from the laboratory line Slab) through eight backcrosses (100–200 virgin females of the Tn line or the Har line crossed with 50–100 Slab males), expected to restore ~96% of Slab nuclear genes. Population cages were then initiated using the Tn and Har backcrossed lines, with an initial frequency of 50% (i.e. 100 males and 100 females Tn were mixed with 100 males and 100 females Har). Three replicates were performed, and all individuals were 1 day old and virgin. Mosquitoes were introduced into the cage at the same time. All population cages employed discrete generations by establishing new cages at each generation using newly emerged adults resulting from the previous generation. Wolbachia infections frequencies in population cages were monitored by PCR assays as described above.
Data analyses
Field data on the geographic distribution of wPip strains in Tunisia were analysed with genepop (Raymond & Rousset 1995; Rousset 2008). Wright's F-statistics (FST) was examined to estimate population differentiation, based on the distribution of genetic polymorphism between populations. All other statistical analyses and modelling were performed using the r software (R Core Team 2013).
Results
A very narrow contact zone between wPip11 and wPip31 Wolbachia strains in Tunisia
We developed a sensitive molecular assay to easily genotype the five wPip groups (Atyame et al. 2011a) and screened Culex pipiens collected as larvae or pupae in Algeria and Tunisia in several sampling campaigns from 1996 to 2009 (Table S1, Supporting information). Mosquitoes from all examined samples were only infected by one of the two molecular wPip strains, referred to as wPip11 and wPip31, and belonging to the wPip groups I and IV, respectively (Duron et al. 2006b; Atyame et al. 2011a). In Tunisia, wPip11 is distributed in the east and south and wPip31 in the north and west, the latter also spread over in Algeria (Fig. 1). Structure analysis of the wPip strains frequency was carried out in two independent groups of sampled localities (Clusters I and II, Fig. 1, separated by a dry area in which no mosquito breeding site could be found) and evidenced, in both groups, spatial autocorrelations in cytotype frequencies (Mantel test on Spearman's rank correlation: b = 0.01, P = 0.001 for Cluster I, and b = 0.01, P = 0.004 for Cluster II). The contact zone was in the centre of northern Tunisia, delineated by the Medjerda River along which most samples were infected by one of either strain (Fig. 1). The transition between wPip11 and wPip31 distribution was sharp, spanning only a few kilometres, for example 3.60 km between Othman (#28, 100% wPip11) and Briss (#38, 100% wPip31). Within the contact zone, 12 localities exhibited a mixture of mosquitoes infected by wPip11 or by wPip31, with a frequency of wPip11 infections ranging from a few percentage (e.g. Zerga, #30), 20–30% (e.g. Fontaine, #50; Aïn Tounga, #51) to 91–97% (e.g. Slouguia, #40; Œufs, #44).

Most wPip31 display unidirectional CI with wPip11; a few show bidirectional CI
We first investigated the CI patterns between wPip11- and wPip31-infected mosquitoes from different areas. We used five wPip11 isofemale lines from three localities and 16 wPip31 lines from eight localities (Table S3, Supporting information). Two isofemale lines isolated from a same locality were used as replicates whenever possible. In all crosses, compatibility or incompatibility was associated with full embryonic viability (HR > 90%) or mortality (HR = 0%), respectively. We never observed CI in crosses between mosquitoes infected with the same wPip strain (data not shown).
We analysed the CI patterns between the wPip11 and wPip31 strains (Tables S3 and S4, Supporting information, about 20 egg rafts per cross). wPip11 males sterilized wPip31 females in all crosses (80 crosses); this was further confirmed by crossing isofemale-derived wPip11 males from three other localities (El Battan, #43; El Manar, #46; and Aïn Tounga, #51) with wPip31 females derived from the Har isofemale line (Harash, #3; Table S5, Supporting information). Most wPip31 males were compatible with wPip11 females (75 of 87 crosses from 20 wPip31 lines; Tables S3B and S4, Supporting information). However, wPip31 males from the Algerian lines Souk1 and Souk2 were incompatible with all wPip11 females assayed (five wPip11 lines tested, Table S3B, Supporting information).
Thus, two strains of wPip31, molecularly indistinguishable for the markers considered, are present in the studied area. The strains where males are respectively compatible or incompatible with the wPip11 females (unidirectional or bidirectional CI) will be thereafter named wPip31_U or wPip31_B (Table 1). Such uni- and bidirectional CIs were also observed between lines from localities where wPip11 and wPip31 strains were sympatric (Zerga, #30; Aïn Tounga, #51; Table S4, Supporting information). Note that all crossed wPip11 lines were compatible among each other, as were the wPip31_U and wPip31_B lines. As no polymorphic markers can discriminate the two wPip31 cytotypes, we deduced their respective frequencies through extensive crossing experiments. We crossed females from the wPip11 isofemale Sok line (Sokra, #37) with wPip31 isofemale-derived males from geographically distant pure wPip31 (Hamra, #29; Ras Rajel, #57; Khetmine, #62) and mixed wPip31/wPip11 (El Manar, #46; Aïn Tounga, #51) populations. We detected the two wPip31 cytotypes in the five locations, whether near or far from the contact zone. The frequency of the wPip31_B cytotype varied from 3% (1/39; Khetmine, #62) to 20% (7/35; Aïn Tounga, #51) with a mean frequency of 12% (Table 2 and Table S6, Supporting information). Thus, the two wPip31 strains are widespread over Tunisia, but wPip31_U is the most frequent.

- 1 = Compatible cross [all hatching rates (HR) > 90%]; 0 = incompatible cross (HR = 0%). Incompatible crosses are bold. wPip11 males are always incompatible with wPip31_U or wPip31_B females, while wPip31_U males were compatible and wPip31_B males incompatible with wPip11 females. So, two crossing types exist between the molecular strains wPip11 and wPip31: unidirectional CI between wPip11 and wPip31_U and bidirectional CI between wPip11 and wPip31_B. Note, however, that the females from wPip31_U and wPip31_B display a similar CI pattern (boxed).
| Localities | Frequency of wPip31 (n) | N | wPip31_U | wPip31_B | Frequency of wPip31_B | 95% C.I. |
|---|---|---|---|---|---|---|
| 29. Hamra | 1 (56) | 56 | 50 | 6 | 0.11 | 0.04, 0.22 |
| 46. El Manar | 0.96 (84) | 38 | 34 | 4 | 0.11 | 0.03, 0.25 |
| 51. Aïn Tounga | 0.90 (40) | 35 | 28 | 7 | 0.20 | 0.08, 0.37 |
| 57. Ras Rajel | 0.98 (66) | 23 | 19 | 4 | 0.17 | 0.05, 0.39 |
| 62. Khetmine | 1 (40) | 39 | 38 | 1 | 0.03 | 0, 0.13 |
| Total | 191 | 169 | 22 | 0.12 | 0.07, 0.17 |
- Estimations were made through analysing the crossing relationships between wPip31 males from isofemale lines isolated from samples collected in five localities and wPip11 females of the isofemale line Sok (from Sokra, #37; see details on Table S6, Supporting information). Numbers preceding the localities are as in Table S1, Supporting information and in Fig. 1. The five localities are located in wPip31 area (i.e. localities with wPip31 frequency >wPip11 frequency). Samples used in crossing experiments were collected in June (#57) and October 2010 (#29, #46, #51 and #62; Table S1, Supporting information). n, total number of mosquitoes examined to estimate the frequency of wPip31; N, the number of wPip31 isofemale lines used in crosses. Confidence intervals (C.I.) were calculated from binomial distribution.
In the field, wPip11 does not outcompete wPip31 as predicted
The situation in which wPip11 males induce incompatibility with wPip31 females, whereas wPip31 males are mostly compatible with wPip11 females, is predicted to evolve towards wPip11 invasion if the host population is panmictic and unstructured. We therefore performed additional samplings in 2010 and 2011 along the contact zone delineated in 2009 and compiled the complete data set over the 7-year follow-up (2005–2011). All localities where the initial frequency of wPip11 or wPip31 was 100% remained stable (Table S1, Supporting information). In all localities where wPip11 was majority, wPip31 frequency was very low and did not vary over time. Remarkably, wPip11 frequency did not change in four localities where wPip31 was majority (Zerga, #30; Dougga, #41; Gaafour, #53; and Utique, #61) and even decreased in three others (El Manar, #46; Aïn Tounga, #51; and Beja Gare, #54). wPip11 frequency increased only in two localities, transiently in Hamra (#29) and more stably in Fontaine (#50; Table S1, Supporting information). Our data thus indicate an overall stability of the contact zone between the molecular strains wPip11 and wPip31 over the 7-year study.
wPip11 outcompetes wPip31 in population cages
This observed stability of the contact zone prompted us to examine which life history traits may interfere with wPip11 invasion. We first tested the invasive capacity of wPip11 in population cages by confronting wPip11- and wPip31_U-infected lines carrying the same nuclear background. Three population cages were initiated using 50/50 as initial frequencies for wPip11 and wPip31_U. Assuming random mating, complete CI, 100% maternal transmission and no differential fitness cost, wPip11 frequency was expected to reach near-fixation (98%) within four generations (Hoffmann et al. 1990). wPip11 frequency increased rapidly and consistently with these expectations (Table 3). The observed wPip11 frequencies were homogeneous between the three cages (Fisher's exact test, P = 0.245) and showed no significant deviation from the expected frequencies (Exact binomial tests on pooled replicates, P = 0.28 and 1 for generations 3 and 4, respectively). This result shows that, in the laboratory, wPip11 invades as expected, suggesting that life history traits other than CI penetrance, Wolbachia transmission, and fitness cost hamper the spreading of wPip11 in the field.
| Generations | Expected frequencies | Observed frequencies (n) | ||||||
|---|---|---|---|---|---|---|---|---|
| Cage 1 | Cage 2 | Cage 3 | ||||||
| wPip11 | wPip31_U | wPip11 | wPip31_U | wPip11 | wPip31_U | wPip11 | wPip31_U | |
| G0 | 0.50 | 0.50 | — | — | — | — | — | — |
| G1 | 0.67 | 0.33 | — | — | — | — | — | — |
| G2 | 0.86 | 0.14 | 0.82 (75) | 0.16 (16) | 0.90 (80) | 0.10 (8) | 0.93 (75) | 0.07 (6) |
| G3 | 0.98 | 0.02 | 0.97 (70) | 0.03 (2) | 1 (76) | 0 (0) | 0.97 (74) | 0.03 (2) |
- Confrontations were performed between the wPip11 strain (from the Tn line) and the wPip31_U strain (from the Har line) introduced into the same nuclear background. Each cage was set up using males (n = 100) and females (n = 100) from the wPip11 line with an equivalent number of males and females from the wPip31_U line. The expected frequencies of wPip11 and wPip31_U strains were estimated assuming no cost and random mating. The number of individuals analysed by PCR assays to measure the frequency of Wolbachia infections frequencies in cages is indicated in brackets (n).
No specific association between wPip strains and Cx. pipiens forms
A straightforward explanation might be that wPip11 and wPip31 differentially infect the pipiens and molestus forms of the Cx. pipiens complex. These two forms show behavioural and physiological differences: the molestus form mates in confined spaces, remains active during winter and can oviposit without a bloodmeal, whereas the pipiens form mates in open spaces, undergoes winter diapause and requires a bloodmeal for oviposition (Fonseca et al. 2004; Farajollahi et al. 2011; Harbach 2012). We genotyped the CQ11 microsatellite loci of 20 mosquitoes infected by wPip11 (Ayed, #24 and Riadh, #27) and 20 mosquitoes infected with wPip31 (Tabarka, #31; Boussalem2, #71; Table S7, Supporting information). We identified the two pipiens and molestus forms and their hybrids (three-level response variable Spec) in either wPip11- or wPip31-infected mosquitoes (two-level variable Inf), in the four populations (four-level variable Pop). No evidence for any preferential association was found [multinomial logit model (Venables & Ripley 2002): Spec = Inf + Pop + Inf.Pop; likelihood ratio test (LRT) for Inf.Pop: χ2 = 0.54, P = 0.76]. Therefore, the stability of the contact zone cannot be explained by differential infection rates of the Cx. pipiens forms.
Assortative mating is insufficient to explain wPip dynamics
Although the kinetics of wPip11 spreading in population cages are compatible with random mating, we further examined the occurrence of assortative mating, known to allow for the stable coexistence of incompatible wPip strains in field populations (Rousset et al. 1991). We first addressed this possibility by confronting one wPip11 isofemale line (Sok from Sokra, #37) with two wPip31_U isofemale lines (Kef1-1 and Kef1-2 from Kef1, #34). Assuming random mating, we expected 25% of infertile egg rafts (Inf) among the offspring [i.e. no hatching (HR = 0%) and 20–80% abortive embryonic development, hallmarks of incompatible crosses]. The random mating hypothesis could not be rejected (Table 4), except in one of the six cages, which produced significantly less Inf than expected (Fisher's exact test, P = 0.02). However, this was not significant after a sequential Bonferroni's correction. We next wished to address the occurrence of assortative mating in the field, by measuring the incidence of sterile egg rafts collected in locations where wPip11 and wPip31 are sympatric (Table 5). Overall, we examined 137–590 egg rafts per locality, that is a total of 1938 egg rafts. In control localities with only wPip11 (Jedaida, #59)- or wPip31 (Hamra, #29 and Dougga, #41)-infected mosquitoes (allopatric), almost all egg rafts were fertile (<1% infertile, Table 5). In localities where both wPip11 and wPip31 were present (Zerga, #30; Oued Melah, #39; El Manar, #46 and Font Mjez, #69, sympatric), the frequency of Inf was between 3% (7 of 252) and 7% (18 of 245). The incidence of Inf in sympatric and allopatric localities (two-level variable Sym) was significantly different (generalized linear model with binomial error Inf = Sym; LRT: P < 10−6), demonstrating that incompatibility is expressed in natural populations.
| Population | Cage | Number of collected egg rafts | Observed frequency of CI egg rafts (n) | P-valuea |
|---|---|---|---|---|
| Sok × Kef1-1 | 1 | 112 | 0.24 (27) | 0.99 |
| 2 | 79 | 0.10 (8) | 0.02b | |
| 3 | 110 | 0.34 (38) | 0.18 | |
| Sok × Kef1-2 | 1 | 142 | 0.23 (32) | 0.78 |
| 2 | 102 | 0.22 (22) | 0.74 | |
| 3 | 74 | 0.22 (16) | 0.85 |
- Each cage was set up using males (n = 100) and females (n = 100) from the Sok line (wPip11) with an equivalent number of males and females from the Kef1-1 or Kef1-2 lines (wPip31_U). The expected frequency of infertile egg rafts was 0.25, assuming unidirectional cytoplasmic incompatibility (CI) and random mating.
- a Fisher's exact test.
- b Significant difference between observed and expected frequencies of infertile egg rafts.
| Infection status of localities | Localities | Year of sampling | N | Frequency of wPip11 | Number of egg rafts examined | Fertile | Infertile |
|---|---|---|---|---|---|---|---|
| wPip11 > wPip31 | 39. Oued Melah | 2009 | 42 | 0.93 | 245 | 227 | 18 |
| 30. Zerga | 2009 | 47 | 0.02 | 252 | 245 | 7 | |
| wPip31 > wPip11 | 46. El Manar | 2010 | 59 | 0.1 | 272 | 257 | 15 |
| 69. Font Mjez | 2010 | 33 | 0.09 | 590 | 573 | 17 | |
| wPip11 | 59. Jedaida | 2010 | 22 | 1 | 157 | 157 | 0 |
| wPip31 | 41. Dougga | 2009 | 23 | 0 | 137 | 136 | 1 |
| wPip31 | 29. Hamra | 2010 | 41 | 0 | 285 | 284 | 1 |
- The infection status, the year of sampling, the frequency of wPip11 estimated by PCR on N mosquitoes, the number of egg rafts collected, and whether the egg rafts hatched (fertile) or not (infertile) are indicated for each locality.
We then estimated the incidence of assortative mating by maximum likelihood: in the absence of assortative mating, the expected frequency of infertile crosses would be PI = p11(1 − p11)(1 + p31B), where p11 is wPip11 frequency and p31B is wPip31_B frequency among wPip31-infected mosquitoes (i.e. crosses between males wPip11 and females wPip31_U, and crosses between wPip11 and wPip31_B in both directions). Assortative mating entails a reduction in this frequency, described as PI (1 − F), where F is the correlation of cytotypes between mates. F and p11 can be jointly estimated using binomial models for the observed frequencies of wPip11 and Inf. However, this gives little information on F when strain frequencies are close to 0 or 1, which is the case for the sympatric populations here, even when a large number of egg rafts are examined. We nevertheless jointly estimated by maximum likelihood (i) the wPip11 frequencies in each population and (ii) a single correlation F, with its confidence interval, for all populations, using p31B = 0.12 (i.e. the observed mean frequency of wPip31_B, Table 2). As expected from strain frequencies near to zero, the confidence interval was very wide [−0.08, 0.63], with an estimated value of F of 0.39.
These results show that wPip11 and wPip31 strains clearly mate at random in cage trials and that CI is expressed in the field where those strains are present. The incidence of Inf in the field remains compatible either with random or with only moderate assortative mating. Therefore, additional factors must be invoked to explain why wPip11 does not supersede wPip31 in Tunisia.
Low dispersal rate and wPip31_B are critical for the stability of the contact zone
The frequency dynamics of the three types can be analysed as that of two types
Only females transmit Wolbachia, and wPip31 females display a similar CI pattern towards wPip11 males when infected by wPip31_B and wPip31_U (Table 1, the rows for wPip31_B and wPip31_U females are identical). This implies that wPip31_U and wPip31_B have the same expected reproductive success and, therefore, that their relative frequency does not change. Thus, the evolution of the molecular polymorphism wPip31/wPip11 can be investigated by models previously developed for two cytoplasmic types (Caspari & Watson 1959; Hoffmann et al. 1990), considering complete CI in one direction and low CI in the other direction, in proportion to the frequency of wPip31_B within wPip31.
wPip31_B initial frequency is a key factor for wPip11 invasion
Analysis of wPip molecular types was carried out in assuming no effect of wPip infections on female fecundity (i.e. no cost of infection) and full maternal transmission of the three cytotypes, as generally observed in Cx. pipiens (Rasgon & Scott 2003). These conditions are well known to lead to an unstable equilibrium, at a frequency depending on wPip31_B relative frequency. If the initial frequency of wPip11 is higher than that of wPip31_B, wPip11 is expected to spread to fixation over a few generations, eliminating both wPip31_U and wPip31_B. If the initial frequency of wPip11 is lower than that of wPip31_B, wPip11 should be eliminated, and wPip31_U and wPip31_B would stably coexist. Hence, in this model, no stable coexistence of the three wPip cytotypes is made possible and wPip11 could colonize localities where wPip31 is present only if its frequency exceeds the frequency of wPip31_B. This suggests that dispersal might be a critical factor for the absence of invasion.
Low dispersal rate may prevent wPip11 invasion
We thus investigated the role of dispersal on the stability of the contact zone by modelling evolution of the spatial frequency cline of the three cytotypes (wPip11, wPip31_U and wPip31_B) along a transect orthogonal to the contact zone. We therefore developed a spatially explicit version of earlier two-patch models (Telschow et al. 2005; Flor et al. 2007). This model (Annex 1) considers the influence of a fecundity (or survival) cost (c) for the wPip11 strain outside its initial range. It assumes that there is no selective difference between wPip31_U and wPip31_B and that their relative frequency remains constant through time. Migration occurs with probability m between adjacent populations across the contact zone. Adjacent populations in the model represent adjacent localities, about 5 km apart, in the habitat. Numerical iterations were performed during 50 generations (i.e. about 5 years). The model allows estimating the threshold value of the dispersal probability m per generation below which wPip11 cannot invade (i.e. it reaches 50% in a population). This dispersal probability was investigated taking into account different values of b (the frequency of wPip31_B) and c (the cost associated with wPip11 infection when outside its initial range). Schematically, m will increase when b or c increases. Even under the least favourable conditions, that is, a relatively high fecundity cost (c = 0.2) and a relatively high wPip31_B frequency (b = 0.16), wPip11 invasion can be halted only if dispersal is extremely low (e.g. a dispersal probability m < 0.08). Given the distance between adjacent populations, this represents a mean square axial dispersal distance σ2 lower than 2 km2 per generation.
Discussion
Our analysis of wPip polymorphism in Culex pipiens field populations from Algeria and Tunisia revealed the presence of two molecular strains, wPip11 and wPip31. These strains are observed in Tunisia with a clear spatial structure: wPip31 over the north and west, and wPip11 over the east and south. Whereas most localities are infected either by wPip31 or by wPip11, they are sympatric along a very narrow contact zone in the centre of the northern part of Tunisia.
We found CI in all crosses between wPip11- and wPip31-infected isofemale lines. In all instances, wPip11 males sterilized wPip31 females. However, while most of the wPip31 males were compatible with wPip11 females, a fraction of wPip31 males, molecularly indistinguishable induced CI (Table 1). Thus, two wPip31 strains, wPip31_U and wPip31_B, coexist in all localities studied. The differences between the two wPip31 strains may be interpreted in terms of different mod factors, as wPip31_U males are incompatible with other wPip strains (Atyame et al. 2011b, 2014).
The main result of this study is the stability of the contact zone observed for over 7 years. Theoretical models predict that, in an unstructured panmictic population, the CI-inducing strain (in the present case wPip11) should invade. We thus explored several mechanisms that could explain why the wPip11 strain does not progress: (i) assortative mating, (ii) local adaptation, (iii) limited dispersal of the wPip11 strain (structuration) and (iv) quality of the breeding sites.
Assortative mating cannot explain the stability of the contact zone
Due to negative effects of CI on host fitness, selection should favour mechanisms that limit or suppress the expression of CI (Rousset et al. 1991; Turelli 1994). Among them, assortative mating between individuals infected with the same Wolbachia may lead to the stable coexistence of incompatible strains of Wolbachia in field populations. Such mating discrimination has been reported between uninfected Drosophila subquinaria females and infected Drosophila recens males when they occur in sympatry (Jaenike et al. 2006). However, we did not detect any preferential association between wPip11 or wPip31 and either Cx. pipiens forms (pipiens or molestus). In addition, we did not detect assortative mating in laboratory trials (Table 4). These results are in line with previous studies showing that Cx. pipiens females cannot discriminate between compatible and incompatible partners (Laven 1967b; Curtis & Adak 1974; Curtis et al. 1982; Duron et al. 2011). Cage population experiments showed that wPip11 rapidly supersedes wPip31_U, as expected from models assuming random mating, complete CI, 100% Wolbachia transmission and no fitness cost. Although the occurrence of moderate assortative mating in the field could not be formally excluded, it would have a limited impact on wPip11 dynamics, even at the maximum value of the confidence interval (Fig. S2, Supporting information). Assortative mating may have a significant impact only if at its highest values and combined with a high infection cost, which is not the case here (see below). Thus, if assortative mating occurs in the field, its incidence is too low to explain the stability of the contact zone.
Differential fitness cost cannot explain the stability of the contact zone
The stability of the contact zone can also be explained by local adaptation. Association between Wolbachia strains and nuclear or mitochondrial genes may indeed confer an increased fitness of mosquitoes infected by a wPip strain at the expense of immigrant mosquitoes infected by another wPip strain. For instance, local adaptation to climate due to mitochondrial genes has already been described (Blier et al. 2001; Ehinger et al. 2002; Fontanillas et al. 2005; Wallace 2007). As mitochondria and Wolbachia are maternally cotransmitted, different Wolbachia strains could thus be strictly associated with different mitochondrial haplotypes that confer differential local adaptation. In our system, we found two mitochondrial cytb haplotypes, each being strictly associated with either wPip11 or wPip31 infections (data not shown), which could potentially participate to local adaptation. However, modelling the cytotype clinal patterns, we showed that wPip11 is expected to invade wPip31 area, even when associated with a very high fitness cost (c = 0.2), providing a dispersal probability over 0.08 (Fig. 2). Such extreme cost is very unlikely, as wPip11 reached near-fixation in population cages as rapidly as predicted by models assuming no differential cost. Moreover, we also compared the observed wPip11 dynamics in the cages with those expected assuming random mating, complete CI, 100% maternal transmission and a differential fitness cost of 0.2: the dynamics was significantly faster (exact binomial tests on pooled replicates, P = 3 × 10−6 and 1 × 10−4, respectively, for generations 3 and 4). Thus, it is unlikely that fitness differences associated with wPip11 and wPip31 could explain the observed stability of contact zone.

Low dispersal likely prevents wPip11 invasion
In a panmictic unstructured population, the coexistence of the three cytotypes (wPip11, wPip31_U and wPip31_B) is not possible. Because wPip11 is favoured by unidirectional CI, wPip31 should obviously be eliminated from localities where wPip11 is the most abundant (i.e. wPip11 > wPip31). wPip11 should also increase up to fixation in localities where wPip31 predominates (i.e. wPip31 > wPip11), when its frequency is higher than wPip31_B. wPip31_B frequency therefore represents the threshold controlling wPip11 invasion. This frequency, estimated in five localities through crossing experiments, ranges from 3% to 20% (mean frequency 12%). Modelling the spatial dynamics of the three cytotypes, assuming a frequency of wPip31_B b = 0.16 and a large cost c = 0.2 of wPip11 (which is unlikely high), we deduced that the mean axial dispersal distance σ2 should be lower than 2 km2 per generation to prevent wPip11 progression from one locality to the next within the 5-year period (about 50 generations). This is much lower than published dispersal estimates in Cx. pipiens, for example σ2 = 43 km2 per generation in southern France (Lenormand et al. 1999). This is within the range of low mean dispersal distances (i.e. 1–2 km) deduced from recent mark–recapture studies in Hawaï (Lapointe 2008), or New York State (Ciota et al. 2012). However, in these studies, traps for recapture were set within 3 and 2 km from the release site, respectively, so that more distant events could not be observed. According to the results of our model, low dispersal currently is the parameter that better explains the contact zone stability. Moreover, this is consistent with previous theoretical studies (Telschow et al. 2005; Flor et al. 2007).
Influence of the quality and/or quantity of breeding sites
Local spatial heterogeneity such as differential host density could also slow down or even block Wolbachia progression (Barton & Turelli 2011). However, wPip11 progression in Tunisia could be stalled only if populations infected with wPip31 are far denser than the nearby populations infected by wPip11. This does not correspond to our field observations, based on the survey of breeding larval sites: most sites were sparsely populated, except for a few along the coast in Bizerte (#15) and Tunis (#16).
These Culex populations may actually be better described as metapopulations with fluctuating local densities, in which case Wolbachia spread could be much slower, as shown by Hancock & Godfray (2012). Mosquito larval and pupal stages require watered breeding sites to develop, and the availability of suitable sites may constitute a limiting parameter. Indeed, Cx. pipiens breeding sites in the contact zone are temporary, dry during the summer due to temperatures above 30 °C and low rainfall, and filled again from autumn to spring. Extinction–recolonization events thus probably reset every year wPip frequencies to those of the breeding adults present when sites are being rewatered. This may act as a brake on wPip11 progression, despite its CI advantage, and may explain why wPip11 did not invade in most of the sympatric sites where its frequency was above 10% (El Manar, #46; Hamra, #29; Aïn Tounga, #51; and Béja Gare, #54) and even in Fontaine (#50) where the wPip11 frequency appears to level-off at 60–70%.
Conclusion
We highlighted the presence of a very narrow contact zone between the distribution areas of the wPip11 and wPip31 molecular strains in Tunisia. The situation is complex due to the segregation of three Wolbachia strains: while the most frequent wPip31 strain is unidirectionally incompatible with wPip11, a wPip31 strain bidirectionally incompatible with wPip11 (wPip31_B) is present at low frequency (~12%). The narrow contact zone between the wPip11 and wPip31 strains appeared stable over a 7-year survey. The situation might have been stable for a longer period of time, as wPip31-infected mosquitoes have been detected in 1996 in Mateur (#14) and Bizerte (#15) and wPip11 in 1997 in Tunis (#16), located 50–60 km apart. The observed stability cannot be explained by local adaptation and probably results from a low dispersal probability strengthened by metapopulation dynamics in Culex pipiens. Although this result has already been predicted theoretically (Telschow et al. 2005; Flor et al. 2007), we present here the first empirical evidence of such a stable coexistence in the field. This study points out that a thorough knowledge of the host dynamics and the environmental conditions prevailing in the studied region is required to understand how Wolbachia distribution evolves in natural populations and how this might impact Wolbachia-based control strategies.
Acknowledgements
This work was performed in collaboration with Comité de Coopération Marseille Provence Méditerranée supervised by Robert Bismuth. We are very grateful to Haoués Alout for samples collected in Algeria; Sandra Unal and Arnaud Berthomieu for technical assistance; Badii Kamoun, Christelle Vidaler, Pierre-Yves Biba Nkouka and Olivier Sacchi for their help in Wolbachia genotyping and crossing experiments; and Pascal Milesi for helpful comments on the manuscript. All sequence data were obtained on the ISEM Environmental Genomic Platform. This study is the contribution 2014-209 of the Institut des Sciences de l'Evolution de Montpellier (Unité Mixte de Recherche, CNRS-IRD-UM2 5554).
Annex 1: Modelling strains dynamics near the contact zone
The evolution of the spatial frequency cline of the cytotypes along a transect orthogonal to the contact zone was modelled. The transect is represented as a linear array of populations. In agreement with data, we assumed panmixia or partial assortative mating of magnitude F, complete transmission and complete CI. Within each site, the change in cytotype i frequency due to incompatibility 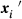 can be represented as
can be represented as 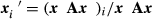 where x is the vector of cytotype frequencies and A is the matrix describing incompatibility, which elements are 1 for compatible crosses and 0 for incompatible ones (see Table 1). We then assumed density regulation, followed by dispersal with a probability m to disperse to the two adjacent populations. In addition, a fecundity (or survival) cost c for the wPip11 cytotype was considered.
where x is the vector of cytotype frequencies and A is the matrix describing incompatibility, which elements are 1 for compatible crosses and 0 for incompatible ones (see Table 1). We then assumed density regulation, followed by dispersal with a probability m to disperse to the two adjacent populations. In addition, a fecundity (or survival) cost c for the wPip11 cytotype was considered.


Numerical iterations of the recursions were performed to check these different conclusions and to further check that local adaptation of cytotypes, in the form of a larger reduction in fecundity of cytotypes outside their initial range (c), did not affect the conclusions. For simplicity, in these computations, edge effects were avoided by assuming a large circular array, with two symmetric clines evolving simultaneously. The initial state of the contact zone was a step function, all wPip11 on one side and all wPip31 on the other. The initial frequency of wPip31_B within all wPip31 was varied around the observed one (i.e. 12%). Dispersal occurred among adjacent populations on the array.
The results of our model are presented in Fig. 2. It displays the maximum amount of dispersal allowed to prevent wPip11 from reaching 50% in a population (i.e. from invading the population) at a given distance from the initial contact zone after 50 generations. It confirms that low dispersal is required to prevent the advance of wPip11 (m < 0.08), even if the infection is associated with a relatively high fecundity cost (e.g. c = 0.2). Although the qualitative conclusions of the analytical approximation are supported, the approximation can substantially overestimate the speed for small dispersal rate among discrete populations (e.g. the approximation overestimates the speed by a factor 2 for m = 0.02 in the absence of any fitness cost). Equivalently, estimates of m deduced from an observed advance using the approximation would be too low. To some extent, this is expected as for low enough dispersal rate—even for weakly bidirectional incompatibility—selection against the rare cytotype will be stronger than immigration, so that there will be no wave of advance (Telschow et al. 2005; Barton & Turelli 2011).

References
C.M.A., O.D., N.P., P.F. and M.W. conceived and designed the experiments. C.M.A., M.B., P.M., O.D., E.D., P.F. and M.W. performed the experiments. C.M.A., P.L., F.R., O.D., N.P., P.F. and M.W. analysed the data. C.M.A., P.L., F.R., N.P., A.B., P.F. and M.W. wrote the article.
Data accessibility
Data sets used in this study are available as Supporting information in the online version of this article.

