

Merging pedigree databases to describe and compare mating practices and gene flow between pedigree dogs in France, Sweden and the UK
Summary
Merging pedigree databases across countries may improve the ability of kennel organizations to monitor genetic variability and health-related issues of pedigree dogs. We used data provided by the Société Centrale Canine (France), Svenska Kennelklubben (Sweden) and the Kennel Club (UK) to study the feasibility of merging pedigree databases across countries and describe breeding practices and international gene flow within the following four breeds: Bullmastiff (BMA), English setter (ESE), Bernese mountain dog (BMD) and Labrador retriever (LBR). After merging the databases, genealogical parameters and founder contributions were calculated according to the birth period, breed and registration country of the dogs. Throughout the investigated period, mating between close relatives, measured as the proportion of inbred individuals (considering only two generations of pedigree), decreased or remained stable, with the exception of LBR in France. Gene flow between countries became more frequent, and the origins of populations within countries became more diverse over time. In conclusion, the potential to reduce inbreeding within purebred dog populations through exchanging breeding animals across countries was confirmed by an improved effective population size when merging populations from different countries.
Introduction
More than 280 Mendelian disorders/traits have been reported in dogs according to the Online Mendelian Inheritance in Animals (OMIA, 2016), with various prevalences and consequences for canine health, and new disorders are added over time. Loss of genetic variability induced by selection on specific characteristics is widely considered to be the cause of the increased observations of inherited disorders in pedigree dogs (APGAW, 2009). Dog breeding is characterized by various breeding practices and regulations across countries (Hedhammar et al. 2011; Shariflou et al. 2011). Hence, it is not only the extensive use of particular animals within a country but also the exchange of breeding animals across countries that can spread inherited disorders in purebred dogs. Additionally, mating between close relatives is known to increase the expression of recessive diseases (Leroy & Baumung 2011). Hence, an improved understanding of genetic variability and potential impacts on health-related issues in purebred dogs is desirable. One tool that might facilitate such an understanding is the ability to share pedigree information across dog registry organizations.
Pooling of pedigrees from different countries has the potential to improve the measurement of genetic variability in target populations and to provide complete information, enabling the avoidance of problematic mating (mating of close relatives or overuse of certain animals) when breeding animals are exchanged internationally. In a study on the Icelandic Sheepdog, Oliehoek et al. (2009) showed that the recent origins of the breed could be traced back to various countries, demonstrating that international exchange of breeding animals has played an important role in the evolution of the breed in recent history. Additionally, Fikse et al. (2013) revealed significant migration of individuals of four pedigree dog breeds between Sweden and the UK, suggesting that pooling of pedigrees from different countries may improve the accuracy of derived indicators of genetic diversity.
We aim to investigate the evolution of inbreeding and breeding practices (mating between close relatives) and to describe gene flow over time by merging pedigrees from several countries (France, Sweden and the UK) for four breeds: Bullmastiff (BMA), English setter (ESE), Bernese mountain dog (BMD) and Labrador retriever (LBR).
Materials
The Société Centrale Canine (France), Svenska Kennelklubben (Sweden) and the Kennel Club (UK) provided pedigree databases for four dog breeds (all records were included and updated to the year 2014) as follows: for BMA, the records included 7877 individuals from France, 4364 from Sweden and 48 176 from the UK; for ESE, the records included 187 247 individuals from France, 12 806 from Sweden and 33 823 from the UK; for BMD, the records included 63 202 individuals from France, 22 569 from Sweden and 28 285 from the UK; and for LBR, the records included 235 141 individuals from France, 112 827 from Sweden and 1 093 630 from the UK.
For each breed, the pedigree databases from the three countries were merged into one combined pedigree database by identifying and replacing IDs for the individual dogs listed with more than one ID. First, we identified duplicates within the pedigree database in each country based on following attributes: dam ID of individual, sire ID of individual, name of individual and birthdate of individual. When the four attributes of two or more individuals were the same, their IDs were regarded as a pair of duplicate IDs. One of the duplicate IDs was retained (preferred ID), and the other(s) were considered to be aliases and removed from the pedigree database. If the removed ID appeared as the dam or sire of other individuals, it was replaced with the corresponding preferred ID. The procedure described above for identifying, removing and replacing duplicate IDs was repeated until no pairs of duplicates could be found in the pedigree database. Afterwards, we merged pedigree databases from the three countries into a combined pedigree database. The procedure for identifying, replacing and removing duplicate IDs across countries was the same as that within countries described above, but we matched the duplicate IDs based on the name of individual and birthdate of individual attributes. Besides, to cope with the incorrect input of characters (often typographical errors), Levenshtein distances (Levenshtein 1966) were calculated to take potential deletions, insertions and reversals of characters into account when comparing attributes. After merging, combined pedigree databases contained 58 789 records for BMA; 217 653 records for ESE; 108 291 records for BMD and 1 426 542 for LBR. In addition, because of duplications between countries, the number of records removed from the pedigree databases was 1173 (BMA), 661 (ESE), 1536 (BMD) and 7312 (LBR).
Methods
After merging the pedigree databases for each of the four breeds, dogs registered in France, Sweden and the UK, born between 1980 and 2014 with both parents known, were defined as the reference population. The reference population was divided into cohorts based on the country of registration (the initial country to assign the dog ID) and year of birth period (intervals of 5 years, from the period 1980–84 to the period 2010–14). To simplify the discussion of the reference populations belonging to different registration countries, we have shortened the label hereafter; for instance, we used ‘French population’ to refer to the reference population of dogs first registered in France.

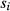 is the relative frequency of use of individual sire i among all sires in a specific reference population. With the exception of NeffS, the other genealogical parameters were computed using PEDIG, a FORTRAN package (Boichard 2002). Coefficients of inbreeding (F) were computed according to the method of Meuwissen & Luo (1992), and coefficients of coancestry (C) were computed following the method of Reynolds et al. (1983). Equivalent complete generations (EqG) were computed as the sum over all known ancestors of the term (1/2t), where t is the ancestor's generation number (Boichard et al. 1997). For a given breed, the effective population size (Ne) was estimated by averaging the increase in coancestry for all pairs of individuals in a reference population (
is the relative frequency of use of individual sire i among all sires in a specific reference population. With the exception of NeffS, the other genealogical parameters were computed using PEDIG, a FORTRAN package (Boichard 2002). Coefficients of inbreeding (F) were computed according to the method of Meuwissen & Luo (1992), and coefficients of coancestry (C) were computed following the method of Reynolds et al. (1983). Equivalent complete generations (EqG) were computed as the sum over all known ancestors of the term (1/2t), where t is the ancestor's generation number (Boichard et al. 1997). For a given breed, the effective population size (Ne) was estimated by averaging the increase in coancestry for all pairs of individuals in a reference population (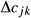 ) (Cervantes et al. 2011):
) (Cervantes et al. 2011):

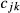 is the inbreeding of a descendent from individuals j and k, and
is the inbreeding of a descendent from individuals j and k, and 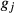 and
and 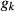 are the discrete equivalent generations for the individuals j and k. Ne was estimated by averaging
are the discrete equivalent generations for the individuals j and k. Ne was estimated by averaging 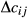 over each generation, using the following formula:
over each generation, using the following formula:

The proportion of inbred individuals was computed as the percentage of individuals with an F higher than 0 when taking into account only two generations of a pedigree according to the method of VanRaden (1992). On the basis of the breed origin of each founder (ancestors without parents known), the founder contributions of different origins were calculated (Boichard et al. 1997). All of the calculated parameters are shown for each cohort of the reference population.
Results
Demographic parameters
The four breeds exhibited different reference population sizes over time in the three registration countries (Table 1). France showed the largest reference populations for ESE (157 186) and BMD (59 848). The UK showed the largest reference populations for BMA (44 377) and LBR (1 037 174). For each breed, the smallest reference population was found in Sweden. The sizes of the French populations increased over time for BMA, ESE and BMD, while decreased beginning in 1995–99 for LBR. The sizes of the Swedish populations started to decrease after 1990–94 for BMA, ESE and BMD, while fluctuated for LBR. The sizes of the UK populations first increased and then decreased for all four breeds.
| Registration country | Birth year period | Bullmastiff | English setter | Bernese mountain dog | Labrador retriever |
|---|---|---|---|---|---|
| France | 1980–84 | 203 | 7643 | 259 | 4529 |
| 1985–99 | 376 | 16 191 | 1468 | 16 835 | |
| 1990–94 | 572 | 22 906 | 4120 | 34 459 | |
| 1995–99 | 883 | 25 874 | 8294 | 48 220 | |
| 2000–04 | 1164 | 26 771 | 11 647 | 44 203 | |
| 2005–09 | 1066 | 28 454 | 16 132 | 33 801 | |
| 2010–14 | 1148 | 29 347 | 17 928 | 35 074 | |
| Sweden | 1980–84 | 102 | 1122 | 1699 | 15 738 |
| 1985–99 | 212 | 1044 | 2024 | 11 724 | |
| 1990–94 | 238 | 1167 | 3042 | 12 177 | |
| 1995–99 | 344 | 1001 | 3035 | 10 526 | |
| 2000–04 | 402 | 947 | 2715 | 12 022 | |
| 2005–09 | 580 | 861 | 2775 | 11 794 | |
| 2010–14 | 332 | 778 | 2307 | 10 694 | |
| UK | 1980–84 | 2035 | 4739 | 1146 | 55 911 |
| 1985–99 | 4337 | 5306 | 3038 | 82 251 | |
| 1990–94 | 6116 | 4678 | 3420 | 127 483 | |
| 1995–99 | 10 936 | 3737 | 4131 | 141 599 | |
| 2000–04 | 9425 | 2746 | 3857 | 190 125 | |
| 2005–09 | 7211 | 1995 | 3500 | 222 462 | |
| 2010–14 | 4317 | 1420 | 2969 | 187 343 |
Both the actual number of sires and NeffS followed the evolution of the size of the reference population for the four breeds in each country (Table S1). The evolution of the ratio of effective number of sires to actual number of sires (NeffS/Number of sires) is presented in Figure S1, where a stable or slightly increasing tendency of more balanced use of breeding animals over time was demonstrated in both the breeds and registration countries.
Genealogical parameters
The amount of pedigree knowledge increased over the period, regardless of the breed and the country of registration (Figure 1). As shown by EqG, pedigree knowledge increased after merging the databases (Table S2). For instance, before merging, the EqG values for BMA in France, Sweden and the UK were 4.15, 3.94 and 8.60, respectively. After merging, the EqG values for BMA in France, Sweden and the UK increased to 4.39, 5.92 and 8.70.

Depending on the period, breed and country, the average F and average C values varied between 0.01 and 0.16 but were below 0.1 in the majority of periods (Figure 2). In general, both the average F and average C values were higher for the UK population than for the French and Swedish populations for all four breeds. In particular, the average C value was below the average F value for the UK population, while these parameters were at the same level in the French and Swedish populations. Additionally, both the average F and average C values slightly decreased or remained stable in all breeds and all countries with the exception of an increase for ESE in the UK and LBR in France and the UK.

The effective population size based on the coefficient of coancestry (Ne) varied from 26 to 425 (Figure 3). Ne decreased for ESE in the UK and LBR in France and the UK over the entire study period, while it increased for the other populations. Throughout the investigated period and for all of the breeds, when we considered the populations from the three registration countries as a single population, Ne was always lower than the sum of Ne for the populations in the three registration countries, indicating a detectable amount of gene flow across countries. However, the global Ne value was always higher than the highest country-specific Ne (considering the three breeds independently), except for LBR. In LBR, global Ne was equivalent to the UK value, probably because the UK subpopulation was demographically much more important than the two others.

Considering two generations, the proportion of inbred individuals ranged from 0.00 to 0.12 (Figure 4). In comparison between the three countries, the proportion of inbred individuals was often highest in the UK populations and lowest in the Swedish populations. Moreover, the proportion of inbred individuals fluctuated for BMA and was stable for other breeds in the French populations, while it tended to decrease in the UK and Swedish populations. Overall, the proportions of inbred individuals, considering the three countries, decreased during the period.

Founder contribution
In Sweden, the origins of populations became more diverse and the founder contributions (from the various origins) became more equal (Figure 5). In France, the population became increasingly diverse over time for BMA; the influence of the dominating origins of founders in 1980–84 reduced considerably for ESE and BMD, and the French LBR population in 1980–84 seems to be substituted through a large influx from the UK. For the native British breeds BMA, ESE and LBR, the founder contributions in the UK populations were dominated by the UK founders, while for native Swiss breed BMD, the founder contributions were dominated by Swiss founders (average 0.49). From 2000–04 onwards, the UK populations showed greater founder contributions from foreign countries than in previous periods. Considering populations from the three countries together, for BMA and LBR, the global founder contributions were still dominated by the UK founders, largely due to the demographic weight of the UK populations. For ESE and BMD, the founder contributions of France and other European countries shifted, but the contribution of the main founder remained fairly stable.

Discussion
The evolution of genetic variability based on the genealogies of purebred dogs has been studied in many breeds (Karjalainen & Ojala 1997; Leroy et al. 2006; Calboli et al. 2008; Leroy et al. 2009; Mäki 2010; Lewis et al. 2015; Wijnrocx et al. 2016). More rarely, studies have considered dog breeds raised in different countries, analysing gene flow between national populations (Oliehoek et al. 2009; Mäki 2010). By analysing four breeds and populations from three different countries, we had the opportunity to investigate the evolution of breeding practices and gene flow over time and across countries.
Merging pedigrees
Merging pedigree databases across countries is complex and time-consuming work that is not confined to dog populations but is also carried out for cattle (Fikse 2004) and horse populations (Viklund et al. 2015). As a prerequisite to explore within-breed genetic variability through international collaborations, an operational method to merge different pedigree databases was needed. We have developed a solution to merge databases from different countries. We observed a substantial improvement in pedigree knowledge, especially for the Swedish population (Table S2).
Two points are worth highlighting based on our experience in handling the pedigree records from four breeds across these three countries. First, the success of detecting duplicates within and between countries relies strongly on the discovery of potential duplicate records (for the same individuals) based on attributes (e.g. name of individual, birthdate of individual) recorded in the databases. However, these attributes may be recorded with missing information, or they may not be exactly the same due to typographical errors, or they may be inconsistently formatted during registration. Second, if a standard format for international dog IDs was proposed and adopted for the registration of pedigree dogs across countries, similar to the Universal Equine Life Number (UELN) system used in horses, the merging of pedigrees would become more efficient and precise.
Comparing genealogical parameters
The results of our study presented herein are similar to other published results for the examined breeds (Leroy et al. 2009; Jansson & Laikre 2014; Lewis et al. 2015). In the French populations, the EqG values for ESE, BMD and LBR born from 2000 to 04 were 6.3, 4.8 and 6.9, respectively, in the present study, similar to results of Leroy et al. (2009), who reported EqG values for ESE, BMD and LBR born from 2001 to 05 of 6.4, 5.0 and 5.5. The average F and average C values for French dogs born from 2000 to 04 differed by 0.012 at most compared with the results for the birth year period of 2001–05 reported by Leroy et al. (2009). For the Swedish populations, the average F values for BMA born from 1990 to 94 and 2005 to 09 were 0.06 and 0.03, respectively, while those for BMD were 0.05 and 0.04 in the present study. These results are similar to those of Jansson & Laikre (2014) for the Swedish population, where the average F values for BMA alive at the end of 1995 and 2010 were 0.06 and 0.03, respectively, while for BMD, they were 0.05 and 0.02. For the UK LBR population, the trends in the average F and average C values observed in our study were the same as those reported by Lewis et al. (2015).
Evolution of breeding practices and gene flow
Mating between close relatives and popular sire effect are considered to be the major causes of overexpression and dissemination of inherited disorders (Leroy & Baumung 2011). When considering mating between close relatives, evaluation of the proportion of inbred individuals, considering two generations of pedigrees in different subpopulations (dogs from the same breed, but registered in different countries), led to interesting results.
In France, a sudden increase in the level of inbreeding occurred in 1995–99 for BMA. This was likely a result of the highest proportion of inbred individuals occurring in 1990–94 (Figure 4). In Sweden, the level of inbreeding and the proportion of inbred individuals has decreased since 1980–85 for BMA, ESE and BMD, while it has been stable or low for LBR (less than 0.02). Moreover, the genetic variability of LBR was not improved according to the level of inbreeding, which may be related to the stability of proportion of inbred individuals in LBR (Figure 2). In the UK, proportions of inbred individuals were the highest among the three countries. Explaining the difference we found between the subpopulations studied is difficult; however, we can link the decrease of this practice in Sweden with the implementation of recommendations aiming at avoiding mating between close relatives due to its potential impact on dog health. Additionally, the average F value was higher than the average C value for the four breeds in the UK, while this was not the case in France and Sweden. In particular, the level of inbreeding and the proportion of inbred individuals has decreased in the UK population from since the 2010s, indicating that breeding practices may have changed. This finding is similar to results published for both vulnerable native breeds and the 20 most common breeds in the UK showing that the mean rate of inbreeding per generation has significantly decreased from 2000 onwards (Lewis et al. 2015). The decreasing proportion of mating between close relatives in Sweden may reflect a more important concern of Swedish citizens towards animal welfare, as illustrated by different surveys (European Commission 2016).
When considering the popular sire effect, it is difficult to clearly determine to what extent the practice has changed over the time period we studied. The actual number of sires and NeffS was mainly related to the evolution of subpopulation size (Table 1). Similarly, the NeffS/Number of Sires ratio was observed to be higher in large subpopulations, indicating less intensive use of sires in large populations. On the other hand, the ratio showed either a stable or increasing trend, even for subpopulations with an increasing population size, which could indicate a tendency towards a more balanced use of sires.
The results of the present study clearly showed that gene flow across countries has become more frequent in all of the countries and all of the breeds we studied. The Swedish populations exhibited the most diverse origins of founder contributions among the three countries based on the four breeds we studied. The UK populations have received a greater founder contribution from overseas, even among native breeds (BMA, ESE and LBR) since 2000–04, which may be related to the relaxation of the UK quarantine law for the importation of dogs (Lewis et al. 2015). Additionally, when gene flow was compared between any two of the countries, Sweden and the UK were found to exhibit the most frequent exchange of breeding animals.
Management of genetic variability
Overall, Ne has increased over the last several years in all of our subpopulations, except for the ESE subpopulation in the UK (which is related to a large decrease in the number of births). This could be related to favourable demographic trends (especially in France), but it could also be related to diversification of origins (through increased gene flow), and possibly to better management of the popular sire effect, although this should be confirmed through more extensive analysis.
The choice of adequate strategies for the management of genetic variability must be made on a breed by breed case in pedigree dogs because each breed faces a unique situation of inbreeding (especially influenced by the evolution of population size). For breeds with a small or decreasing population size, cross-breeding (or introgression) can be an interesting solution to reduce inbreeding or ensure genetic improvement (Pirault et al. 2013; Leroy et al. 2014). However, cross-breeding is rarely practised in pedigree dogs, which makes the management of genetic variability within a breed more challenging. Improvement of within subpopulation genetic variability could be achieved by exchanging breeding animals between countries. This is illustrated by the observation that, in the breeds with a small or medium population size that we studied (BMA, ESE and BMD), Ne was always higher when considering the dogs registered in all three countries together than when considering only a single country. This result illustrates the value of genetic exchanges between subpopulations, as is theoretically well established (Wright 1931). Conversely, for breeds with a large population size, such as LBR, Ne was not higher when considering the dogs registered in all three countries together than in the French population alone and was only a slightly higher than in the UK population. It appears that the effect of gene flow was ‘diluted’ within this breed with a large actual population size. At a more global level, an equal use of sires, but restricted number of litters, could be a solution to improve the effective population size (Windig & Oldenbroek 2015).
Considering the registration of dogs without pedigree but with appropriate phenotype (or even genotype), could constitute another interesting option to increase the breeding population, and it opens up for the possibility to register dogs with different genetic background, which could be assessed through adequate genetic testing. As an example, registries in France are open for most breeds (only 15 breeds have their pedigree closed), meaning that in theory, a dog without pedigree can be registered if it is judged to have the adequate phenotype. However, in practice, those registrations represented in 2014 were only 0.4% of the total number of registration for the corresponding breeds. Therefore, this approach could be efficient only if breed clubs were much less strict in the acceptance of dogs without pedigree. Optimal contribution selection is another solution for managing genetic variability that is potentially more efficient but also results in greater constraints for breeders. Future research could focus on how to fairly compare the genealogical parameters of potential breeding animals across countries, and monitor the consequence on genetic variability and management of inherited diseases, simple or more complex (e.g. hip dysplasia), at an international scale.
Conclusion
Merging pedigree databases across countries and quantifying breeding practices and gene flow for pedigree dogs is possible. Pedigree knowledge was improved after merging pedigrees in the four breeds studied. The breeding practice of mating between close relatives varied between countries for the four breeds. Moreover, a clear tendency of increased gene flow between countries over time was shown for all of the breeds and countries. More importantly, the potential to improve genetic variability within purebred dogs through exchanging breeding animals across countries was confirmed by an improved effective population size when merging the populations of different countries.
Acknowledgements
Shizhi Wang benefited from a joint grant from the European Commission within the framework of the Erasmus Mundus joint doctorate ‘EGS-ABG’. We are grateful to EGS-ABG for funding and grateful to the Société Centrale Canine (France), Svenska Kennelklubben (Sweden) and the Kennel Club (UK) for funding and data provision. We are grateful to the comments on the manuscript of Etienne Verrier, Åsa Viklund and Claire Wade. In addition, we are appreciated of the help from Claire Wade for English language reviewing.

