Concurrent visuomotor behaviour improves form discrimination in a patient with visual form agnosia
Abstract
It is now well established that the visual brain is divided into two visual streams, the ventral and the dorsal stream. Milner and Goodale have suggested that the ventral stream is dedicated for processing vision for perception and the dorsal stream vision for action [A.D. Milner & M.A. Goodale (1995) The Visual Brain in Action, Oxford University Press, Oxford]. However, it is possible that ongoing processes in the visuomotor stream will nevertheless have an effect on perceptual processes. This possibility was examined in the present study. We have examined the visual form-discrimination performance of the form-agnosic patient D.F. with and without a concurrent visuomotor task, and found that her performance was significantly improved in the former condition. This suggests that the visuomotor behaviour provides cues that enhance her ability to recognize the form of the target object. In control experiments we have ruled out proprioceptive and efferent cues, and therefore propose that D.F. can, to a significant degree, access the object's visuomotor representation in the dorsal stream. Moreover, we show that the grasping-induced perceptual improvement disappears if the target objects only differ with respect to their shape but not their width. This suggests that shape information per se is not used for this grasping task.
Introduction
It is now well established that the visual areas of the cerebral cortex are divided into two visual streams: the ventral and the dorsal stream (Ungerleider & Mishkin, 1982; Van Essen, 2005). According to Milner & Goodale (1995), the ventral stream processes visual information for perceptual purposes. The dorsal stream in contrast uses visual information to guide action (visuomotor stream). This distinction is based largely on studies with the form-agnosic patient D.F. (Goodale & Milner, 2004). She suffered extensive lesions to her ventral stream (Milner et al., 1991; James et al., 2003), which left her almost unable to discriminate visual forms (Milner et al., 1991). However, in grasping studies she showed an implicit ability to adjust her grasp aperture to the different forms of visually presented objects (Goodale et al., 1991). These and similar observations provided support for the two-visual stream framework (Goodale & Milner, 1992, 2004; Jeannerod & Rossetti, 1993).
One aspect of D.F.'s behaviour, which is puzzling in this context, is her apparent inability to use the visual information available to her visuomotor stream directly to improve her performance in perceptual tasks. It is obvious that her performance in perceptual tasks is worse than her performance in visuomotor tasks (Goodale et al., 1991; Milner et al., 1991), but the reason for this is not clear. One explanation would be that the two streams are not connected. However, there is evidence that such anatomical connections do exist (Felleman & Van Essen, 1991). A second explanation might be to assume that D.F.'s lesion also destroyed these connections.
A third possible explanation could be that the relevant information in the dorsal stream is only available in the context of a concurrent visuomotor task. If that were true, we would predict that D.F.'s shape-discrimination performance should improve in the context of a relevant visuomotor task. This prediction was tested in a study by Murphy et al. (1996). They found that D.F.'s shape discrimination did improve during a grasping task, but the possibility that proprioceptive or efferent cues rather than visual cues helped D.F. in this condition could not be excluded, and indeed were proposed as the explanation. We revisited this question but included conditions that allowed us to compare the contributions of proprioceptive, efferent and visual cues to D.F.'s shape-discrimination performance in the context of visuomotor tasks. We also examined whether D.F. relied on form or size information by including a condition where only form but no size cues were available.
Our rationale for performing these experiments was that by studying D.F., we might be able to uncover interactions between the two visual streams that would not be measurable in healthy subjects, whose shape recognition is at ceiling. In D.F., the higher levels of the ventral stream have been deprived of their normal input from earlier shape areas, due to the severe damage sustained by the lateral occipital area (LO). Our hypothesis was that D.F. might be able to gain access to the products of object processing in the dorsal stream.
Materials and methods
Subjects
Patient D.F. suffered a carbon monoxide intoxication in 1988 that led to extensive bilateral brain damage (Milner et al., 1991). During a recent magnetic resonance imaging (MRI) examination it was found that her brain lesions are mostly confined to the ventral lateral occipital cortex, comprising in both hemispheres parts of Brodmann areas 18 and 19, but largely excluding V1 and the fusiform gyrus (James et al., 2003). The site of her lesions match very well the site of area LO (Malach et al., 1995; Kanwisher et al., 1996; Kourtzi & Kanwisher, 2000; Grill-Spector et al., 2001). Area LO has been identified by fMRI studies as important for visual object recognition (James et al., 2000; Kourtzi et al., 2003; Stanley & Rubin, 2003; Ferber et al., 2005). D.F. suffers from visual form agnosia, i.e. she has trouble discriminating between different visual shapes, forms, orientations and distances. Apart from that her perceptual abilities are largely intact, e.g. she has normal visual acuity and relatively good luminance, colour and texture perception (Milner et al., 1991). At the time of testing she was 50 years old. Five healthy, age-matched women (mean age: 51.2 years, SD: 7.7) served as control subjects. All subjects were right-handed by self-report. All experiments were undertaken with the understanding and written consent of each subject. The study conforms with the Code of Ethics of the World Medical Association (Declaration of Helsinki), as printed in the British Medical Journal (18 July 1964) approved by the Ethics committee of the Department of Psychology, Durham University.
Experimental procedures
Five experiments were carried out. Common to all experiments was a shape-discrimination task, i.e. subjects had to report verbally whether the target object was a square or a rectangle. An overview of the five different experiments is presented in Table 1. The setup for the experiments is illustrated in Fig. 1.
| Event | Verbal response | Motor response | Objects | |
|---|---|---|---|---|
| Experiment 1 | ||||
| (1) Start of trial: | Shutterglasses open | Occurs at any timeafter start of trial | None | Shapes havedifferent width |
| (2) After 2 s delay: | Shutterglasses close | |||
| Experiment 2 | ||||
| (1) Start of trial: | Shutterglasses open | Occurs at any timeafter start of movement | Grasping with right hand | Shapes havedifferent width |
| (2) Onset of motor response: | Shutterglasses close | |||
| (3) Photosensor blocked: | Object moves out of reach(only in case of non-contact trials) | |||
| Experiment 3 | ||||
| (1) Start of trial: | Shutterglasses open | Shapes havedifferent width | ||
| (2) After 1 s delay: | ‘Beep’ prompts verbal response | Occurs after 1st beep | ||
| (3) Further delay of 1 s: | A 2nd‘Beep’ prompts motor response | Grasping with right,left or both hands | ||
| (4) Onset of motor response: | Shutterglasses close | |||
| Experiment 4 | ||||
| (1) Start of trial: | Shutterglasses open | Occurs at any timeafter start of movement | Pointing with right hand | Shapes havedifferent width |
| (2) Onset of motor response: | Shutterglasses close | |||
| (3) Photosensor blocked: | Object moves out of reach(only in case of non-contact trials) | |||
| Experiment 5 | ||||
| (1) Start of trial: | Shutterglasses open | Occurs at any timeafter start of movement | Grasping with right hand | Shapes haveidentical width |
| (2) Onset of motor response: | Shutterglasses close | |||
| (3) Photosensor blocked: | Object moves out of reach(only in case of non-contact trials) | |||

Illustration of the setup used for the five experiments. At the beginning of the trial the subject's hand rested on the start switch. The release of the start switch indicated the start of the subject's movement. In some trials (non-contact trials) an escape movement of the target object was triggered as soon as the subject's hand crossed the line between the two photosensors.
The first experiment served as the baseline task for the other experiments. It was a simple shape-discrimination task. No concurrent motor task was used. Subjects were wearing LC shutter glasses (Plato, Toronto, Canada), which were closed before the beginning of each trial, and opened at the start of the trial. The LC shutter glasses remained open for 2 s, and remained closed until the start of the next trial. The target objects were presented standing upright at a distance of 50 cm. The square object had a width of 6.50 cm, and the rectangular object had a width of 7.27 and height of 5.81 cm, corresponding to an aspect ratio of 80%. The depth of all objects was 1.5 cm. The surface area of both objects was identical. These objects were chosen because previous experiments with D.F. had shown that she was unable to discriminate between them. The purpose of Experiment 1 was to confirm these earlier findings. The square was used for half of the trials; different trials were intermingled in a random sequence; a total of 130 trials was presented.
Experiment 2 combined the shape-discrimination task with a grasping movement. Subjects had to grasp the target object with their right hand and call out the shape of the object during the movement. Subjects were asked to use a precision grip, i.e. to pick up the object with thumb and index finger. The sequence of events was similar to that in Experiment 1. The LC shutter glasses opened at the start of the trial, and closed at the onset of the hand movement, i.e. the release of the start button. In some of the trials the target object was moved out of reach before the subject could contact the object. This was done using a system developed for catching studies (Schenk et al., 2000). This system uses two motor-driven linear axes moving a sledge that carries a magnet. The sledge can move along any trajectory along the horizontal plane. The axes and the sledge are covered by a steel plate. The target object sits on a magnetic carrier on top of the steel plate, and thus follows the movements of the magnet below the steel plate. This system is connected to a pair of photosensors. In our experiments, the photosensors were triggered when the subject's hand came within 20 cm of the target object, the signal from the photosensors then triggering the onset of object motion, thus putting the target object beyond the subject's reach. For this purpose the object was accelerated at 7 m/s2 for 25 cm, reaching a halt at a distance of 1 m. These trials were called ‘non-contact’ trials and were introduced to ensure that subjects could not use haptic information to guide their decision on the shape of the target object. Only results from the non-contact trials were used in our analysis. These trials were intermingled with contact trials, where the object did not move, and subjects were thus able to contact and pick up the object. Half of the trials were non-contact trials. In the case of both contact and non-contact trials, half of the trials used the square objects. A total of 260 trials was presented to the subjects in this experiment. The hand movement was measured using a 3D movement registration system (Fa. Zebris, Tuebingen, Germany). Two markers were attached to the nails of the index finger and thumb of the right hand. Data were recorded with a sampling frequency of 50 Hz.
In Experiment 3 we tested the possibility that proprioceptive or efferent information from the grasping movement might be used to facilitate the shape-discrimination performance. In particular we were interested in the contribution of proprioceptive or efferent information from the peak hand aperture, because this is the parameter of the grasping movement most closely linked to the shape of the target object (Paulignan et al., 1991; Rosenbaum, 1991; Hoff & Arbib, 1993; Jeannerod, 1996; Smeets & Brenner, 1999). Proprioceptive information was excluded by asking subjects to call out the shape just before the start of their movement. This was achieved by a series of tones that were identical in duration and frequency. The first tone, which occurred 1 s after the start of the trial, prompted the subject to name the shape of the object, and the second tone, which occurred after another 1 s, prompted the subject to start the hand movement. Trials where the subject either named the object after the second tone, or started the movement before the second tone, were discarded and repeated in random order at the end of the experiment. Because the shape-discrimination response in this experiment was concluded before the start of the hand movement, any haptic information about the shape that the subject might have obtained would have come too late to influence the discrimination response. For this reason there was no need for non-contact trials in this experiment. The role of efferent cues was addressed by comparing the accuracy of D.F.'s verbal report and her grip adjustment. We assumed that if efference information guided her verbal report then there should be a close link between the accuracy of her report and the accuracy of her motor response (i.e. the peak grip aperture). To manipulate the accuracy of the motor response we asked D.F. and the other subjects to use different hands for different trials. Subjects were informed before the start of the trial that they should either use the right, left or both hands. In a third of the trials the right hand was to be used, in another third the left, and in the remaining trials subjects were asked to use the index fingers of the two hands together to pick up the object. To measure the movements of both hands, four markers were attached to the index finger and thumb of both hands. A total of 390 trials was performed by the subjects in this experiment.
Experiment 4 was designed to explore the relationship between the motor task and the improvement in perceptual performance. In particular, this experiment addressed the question of whether a non-specific activity-related attentional enhancement might be sufficient to explain the increased perceptual improvement observed in Experiments 2 and 3. If this explanation was correct, we would expect that any concurrent motor activity would lead to improved shape discrimination. A pointing task was chosen because, although very similar to the grasping task, it is different in that size and shape of the object are not strictly relevant for performance of the task. In all other respects this experiment was identical to Experiment 2.
Experiment 5 is a variation of Experiment 2 (i.e. a combination of a shape-discrimination and a grasping task). In this experiment we examined which specific aspect of the target object is used as a cue for the shape-discrimination task. In Experiments 2 and 3, shape and size information were confounded. In Experiment 5, these two aspects were disentangled by using target objects of identical width, but different shape. In this case size or width could not be used to discriminate between the shapes. Two pairs of objects were used. The first pair of objects had a width of 7.27 cm; the second pair of objects had a width of 8.16 cm. Each pair comprised a rectangle with a width : height ratio of 5 : 4 (using heights of 5.81 cm and 6.52 cm in the first and second pair, respectively) and a square (i.e. height and width identical). To obtain equal numbers of trials for each specific object, we used 32 trials per object and trial type (i.e. non-contact vs. contact trials), leading to 128 non-contact trials, and thus 256 trials in total.
Each subject took part in three sessions. Each session lasted for approximately 90 min with a break of 10 min after the first 45 min. The five experiments were repeated in each session in a different sequence.
Analysis and statistics
Because control subjects performed the shape-discrimination task in all five experiments without error, only D.F.'s results were subjected to a formal analysis. As is the case in the analysis of most single-case studies, trials were treated as cases. Thus, the sample of cases that could be used for each experiment ranged between 128 and 130 trials, which means that a normal distribution could be assumed for the statistical analysis. In general, we adopted a significance threshold of 5%; however, in the case of multiple comparisons, we used a Bonferroni correction (when available) or a stricter threshold of 1%.
For the analysis of D.F.'s verbal responses, binomial tests were used to check whether the number of correct verbal responses differed significantly from chance. To compare proportions of correct responses across tasks we used χ2 tests.
For the analysis of D.F.'s grasping response in Experiments 2 and 3, we first smoothed the kinematic traces using a non-parametric Kernel filter (Marquardt & Mai, 1994) and then determined the peak grip aperture for each trial in each hand condition. Grip aperture refers to the 3D distance between the two digits that were used in the grasp (namely index and thumb in the case of unimanual graSPS, or index fingers of the two hands in the case of bimanual graSPS). We used a two-factor between-subject anova to examine the effects of object shape (i.e. square vs. rectangle) and hand (i.e. right, left, both) on maximum grip aperture. Independent-sample t-tests were used for specific comparisons.
To compare the verbal and the grasping response in Experiments 2 and 3, we first transformed the values of the two response types into a common format. For this purpose, d′ values were computed for both response types. The parameter d′ corresponds to the distance between the z-transformed distribution of the responses for two different categories (i.e. in our case square vs. rectangle). It is thus a measure of the accuracy with which it is possible to discriminate between the two categories on the basis of the obtained responses (Gescheider, 1997: see pp. 116–124). To obtain d′ values for both response types we used the formulae (5.3–5.6) provided by Gescheider (1997: pp. 118–119). In the case of the discriminative (verbal) response, d′ was computed on the basis of the proportion of correct and incorrect responses, while in the case of the motor response the means of the peak grip aperture for squares and rectangles, and their standard deviations, were used to calculate d′.
Although d′ is the conventional parameter used to measure and compare discrimination performance, it has the disadvantage that only one value per condition is computed, and thus no statistical comparison is possible. Therefore, as an alternative approach to comparing verbal and motor responses, we also computed expected proportions of correct responses on the basis of the motor response. This allowed us to employ χ2 tests to test whether the actually obtained proportion of correct responses differed significantly from those proportions that were expected if D.F. had based her response on the maximum grip aperture used during the grasping response. For the calculation of the expected proportion, we assumed that D.F. would need to adopt a criterion value for the peak grip aperture and then compare the maximum grip aperture in each trial to decide whether this response was targeted at the square (maximum grip aperture below the criterion value) or the rectangle (maximum grip aperture above the criterion value). In order to make the comparison between verbal and motor responses as conservative as possible, we assumed that D.F. would have used the optimal criterion (i.e. the criterion that would have yielded the highest number of correct responses). Both the d′ calculations and the computation of expected proportions were conducted for all three hand conditions.
Results
It should be noted that all control subjects performed the shape-discrimination task without error in all five experiments. Accordingly we will restrict our presentation of the results to those obtained in patient D.F.
Experiment 1
D.F.'s ability to discriminate between a square and a rectangle was at chance in this purely perceptual task (49.2%, Fig. 2).

Shape-discrimination performance (by verbal report) in Experiments 1–5. The bars depict the proportion of trials where D.F. recognized the shape of the target object correctly. The solid line indicates the values expected in the case of chance performance. D.F.'s shape discrimination was only better than chance in Experiments 2 and 3. The results from Experiment 3 are restricted to trials where the right hand was used. Control subjects performed without error in all five experiments. **P < 0.01.
Experiment 2: verbal performance
In this experiment, she was asked to name the shape while she was reaching forward to pick up the object. In this condition her ability to recognize the shape improved significantly above chance (75%, P < 0.0001, Fig. 2).
Relation between motor and verbal performance
To compare D.F.'s motor and verbal performance we used two approaches. First, we transformed both the grip aperture and the results from the verbal report into d′ measures. We found that d′ (d′ = 1.34) based on her verbal report was substantially higher than that based on her grip apertures (d′ = 0.58). Secondly, we also computed the proportion of correct responses that would be expected if D.F. had used the width of her maximal grip aperture to determine the shape of the object. Again, we found that the number of correct responses that was actually obtained in her verbal report (75%) was significantly higher (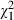 = 4.56; P = 0.033) than the proportion of correct responses that was expected on the basis of the distribution of her grip apertures (62%). In summary, this suggests that proprioceptive or efferent information about the grip aperture cannot explain the high accuracy of D.F.'s verbal discrimination performance in this condition.
= 4.56; P = 0.033) than the proportion of correct responses that was expected on the basis of the distribution of her grip apertures (62%). In summary, this suggests that proprioceptive or efferent information about the grip aperture cannot explain the high accuracy of D.F.'s verbal discrimination performance in this condition.
Experiment 3
The purpose of this experiment was to test whether proprioceptive or efferent information about the peak grip aperture used for a specific object could explain why D.F.'s shape-discrimination performance was better during the grasping task. To explore the role of proprioceptive information, D.F.'s shape-discrimination performance in Experiment 3 (no proprioceptive information available) was compared with her discriminative performance in Experiment 2 (proprioceptive information available). To explore the role of efferent information, we examined the relationship between the accuracy of D.F.'s verbal report on the shape and the accuracy with which she adjusted her grip aperture to the differently shaped objects in the three different hand conditions (right, left, both hands). We will first describe her performance on the verbal shape-discrimination task, and then present the findings on grip aperture. Finally, we will examine how the differences in motor performance obtained for the three different hand conditions relate to D.F.'s verbal performance in those conditions.
Verbal performance
D.F.'s shape-discrimination ability was significantly above chance irrespective of the hand with which she tried to grasp the object (P < 0.0001 for right, left and both hands). Of particular importance is her performance in the right-hand condition. This performance can be compared directly with her performance in Experiment 2, where it was also the right hand that was used. However, in contrast to Experiment 2, where proprioceptive information was generally available, this information was unavailable in Experiment 3 because the verbal response was given before the start of the movement. Nevertheless, D.F.'s shape-discrimination performance was not significantly different in the two experiments (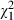 = 0.52; P > 0.40, Fig. 2). This suggests that D.F. did not use proprioceptive information. The finding of above-chance shape discrimination in this condition also seems to suggest that the planning of a grasping movement is sufficient to trigger an improved perceptual performance. With respect to the question of whether D.F. might have used efferent information, it is interesting to note that her discrimination performance is worst in the bimanual condition (see Fig. 5). However, this is the very condition in which her grip is most differentiated for different object shapes (cf. 3, 4). This suggests that there is a poor correlation between D.F.'s verbal and motor performance, and thus little support for the idea that she used efferent information for the shape-discrimination task. This will be explored further by means of a formal comparison of D.F.'s motor and verbal performance (see below).
= 0.52; P > 0.40, Fig. 2). This suggests that D.F. did not use proprioceptive information. The finding of above-chance shape discrimination in this condition also seems to suggest that the planning of a grasping movement is sufficient to trigger an improved perceptual performance. With respect to the question of whether D.F. might have used efferent information, it is interesting to note that her discrimination performance is worst in the bimanual condition (see Fig. 5). However, this is the very condition in which her grip is most differentiated for different object shapes (cf. 3, 4). This suggests that there is a poor correlation between D.F.'s verbal and motor performance, and thus little support for the idea that she used efferent information for the shape-discrimination task. This will be explored further by means of a formal comparison of D.F.'s motor and verbal performance (see below).

A comparison of D.F.'s discriminative and motor responses in Experiment 3 using two different measures of accuracy. (A) d′ measures the accuracy with which the two shapes are discriminated. Empty bars depict d′ based on the motor response (i.e. maximum grip aperture); filled bars show d′ based on the verbal response (i.e. proportion of correct responses). As measured by d′, discrimination accuracy is consistently (and in the case of the unimanual conditions substantially) higher than the d′ for the motor response. (B) Empty bars depict the proportion of correct shape-discrimination responses that would be expected if D.F. had based her verbal response on the maximum grip aperture attained in any given trial. Filled bars depict the actual proportion of correct responses that were obtained in the verbal report. In the case of the unimanual conditions, the actual proportion of correct responses is significantly higher than the proportion that would be expected on the basis of the motor response. *P < 0.05 and **P < 0.01.

Averaged time-course of D.F.'s grip apertures in Experiment 3. Trajectories for each hand (left hand, both hands, right hand) are presented in separate graphs. The mean trajectory for trials with the square and the rectangle are represented by separate curves. The grasp trajectories for the two different shapes only differ in the case of bimanual grasping.

Mean of D.F.'s maximum grip aperture in Experiment 3. The different hands are presented in different groups. Empty bars represent the mean (and standard deviation) of trials with the square; filled bars represent trials with the rectangle (*P < 0.05 and **P < 0.01). Using the more stringent criterion of %, a significant difference (P < 0.01) between the grip aperture for the square and the rectangle is only found in the case of bimanual grasping.
Peak grip aperture
To examine the effect of hand and shape on peak grip aperture, an anova with hand (right, left, both hands) and shape (square vs. rectangle) as between-subject factors was carried out. Both factors, shape (F1,384 = 19.98; P < 0.0001) and hand (F2,348 = 715.14; P < 0.0001) produced significant main effects. The effect of shape reflects a slightly bigger grip aperture for the rectangle (139 mm) than for the square (134 mm). The effect of hand is produced by a significantly bigger grip aperture for the bimanual condition (162 mm) than for both the right hand (124 mm) and the left hand condition (123 mm). Interestingly, we also obtained a significant interaction between shape and hand. This interaction effect can be explained by the fact that a significant difference between the square and rectangle condition is only obtained in the bimanual condition (t128 = 4.375; P < 0.001). In the right-hand condition the difference approached significance, but did not satisfy the more stringent significance criteria of 1% adopted for post hoc comparisons. In the left-hand condition, the peak grip aperture for the square and rectangle were not significantly different from each other (t128 = 0.032; P > 0.50). These results are illustrated in 3, 4.
Relation between motor and verbal performance
Again we computed d′ and the expected proportion of correct responses on the basis of the obtained distribution of grip apertures to compare D.F.'s motor and verbal performance in this experiment. As can be seen in Fig. 5A, d′ based on the verbal report is consistently higher than that based on grip aperture. This indicates that the shape-discrimination performance obtained in the verbal report is higher than what could be expected purely on the basis of the grip aperture information. Secondly, we also computed the proportion of correct responses that would be expected if D.F. had used the width of her maximal grip aperture to determine the shape of the object. Again, we found that the number of correct responses that would be expected on this basis was, at least in two out of three conditions, significantly smaller than the proportion of correct responses that were actually obtained (right hand: expected proportion: 60%; actual proportion: 78%; 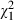 = 9.53; P = 0.0021; left hand: expected proportion: 58%; actual proportion: 71%;
= 9.53; P = 0.0021; left hand: expected proportion: 58%; actual proportion: 71%; 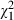 = 4.83; P = 0.028; see Fig. 5B). In fact, in the case of left-hand trials we would predict that on the basis of D.F.'s grip aperture values, her shape-discrimination ability would not be significantly different from chance, when in fact her performance was clearly above chance (P < 0.0001). Taken together, our findings suggest that D.F.'s superior shape discrimination in association with grasping cannot be explained by her use of proprioceptive or efferent cues from her grasping responses.
= 4.83; P = 0.028; see Fig. 5B). In fact, in the case of left-hand trials we would predict that on the basis of D.F.'s grip aperture values, her shape-discrimination ability would not be significantly different from chance, when in fact her performance was clearly above chance (P < 0.0001). Taken together, our findings suggest that D.F.'s superior shape discrimination in association with grasping cannot be explained by her use of proprioceptive or efferent cues from her grasping responses.
Experiment 4
To test whether D.F.'s improvement of shape discrimination found during the grasping task was specific to grasping, we repeated Experiment 2 in a modified version, asking her to point to, rather than grasp, the target object. In this case her discrimination performance was not significantly above chance (P = 0.188). The essential difference between the grasping and the pointing task is that while aspects of the object's shape, namely its width, have to be processed for grasping, they are not needed for pointing.
Experiment 5
Given that during grasping it is actually the object's width and not its shape per se that is used to guide the grasping response, we examined whether D.F.'s ability to discriminate shapes would also improve in a condition where the grasping movement is directed towards objects of different shape but identical width. It turned out that in this condition D.F.'s ability to discriminate between the two shapes was at chance (P = 0.331).
Discussion
Our results indicate that the dorsal representation of visual object information in patient D.F. can influence her ability to distinguish shapes. This is implied by the finding that D.F.'s ability to discriminate different shapes is significantly improved in the context of a grasping task. This finding suggests that during the grasping task information about the shape that was previously inaccessible to D.F. becomes now available. However, before we come to the conclusion that it is visuomotor information from the dorsal stream that supports this improvement, we need to rule out other possible sources of information that also become available during the grasping movement and might have helped in identifying the shape of the target objects. These additional sources of information are proprioceptive and efferent cues.
The results from Experiment 3 indicate that neither proprioceptive nor efferent cues can explain D.F.'s superior shape-discrimination performance during the grasping task. The relevance of proprioceptive cues can be ruled out on the basis of the finding that D.F.'s ability to discriminate between shapes is not diminished in a condition where she has to name the shape first and move afterwards. In this condition, proprioceptive information is not available during the shape-discrimination task. Nevertheless, D.F. might have already planned the grasping movement at that stage, and this means that in principle efferent information from the grasp might have been available. However, because the efferent information is just a copy of the motor commands that actually determine the grasping movement, one might expect that the information contained within the efferent copy is closely related to the characteristics of the actual formation of the grip. Accordingly, it would be expected that in this case the accuracy of D.F.'s verbal report would be closely related to the accuracy of her grip formation. However, D.F.'s verbal accuracy in recognizing the shape of the object was consistently and significantly higher than the accuracy of her grip formation. This dissociation was most striking in the case of left-hand grasping. Here her grip apertures when reaching for the square and the rectangle were practically identical, but her verbal report nevertheless showed that she could quite reliably discriminate between the two objects. In conclusion, these findings suggest that D.F.'s improved shape-discrimination performance during the grasping task was based neither on proprioceptive nor on efferent information. This conclusion is quite different from that of previous studies, which examined the effect of movement cues on the perceptual performance of patients with visual form agnosia. In those earlier studies, proprioceptive and efferent cues could not be ruled out, and therefore the observed perceptual improvements were always attributable to the patient's use of non-visual motor cues (Goldstein & Gelb, 1918; Landis et al., 1982; Murphy et al., 1996; Dijkerman & Milner, 1997).
There is one further alternative explanation, which is worth considering, namely that the improvements observed during the grasping task are induced by attentional changes. It is well established that motor activity or even action-related visual stimuli can improve performance of patients with attentional deficits, most notably in patients with spatial neglect (Robertson et al., 1995; Humphreys & Riddoch, 2001; Humphreys, 2004). It should be noted that this account would need to assume that the attentional effects are fairly specific, otherwise it could not explain why other motor tasks [i.e. pointing (cf. Experiment 4) or the grasping of objects of different shapes but identical width (cf. Experiment 5)] did not also yield improvements in perceptual performance. Nevertheless, specific attention-enhancing effects of motor activity could explain D.F.'s improved performance in the grasping task. However, as has been said many times before, attention and representation are not independent from each other. Attention must act on a given representation, and thus the question arises as to which representation is enhanced by the concurrent motor activity. If we assume that it is the visuomotor processing in the dorsal pathway that is enhanced, we end up with an explanation that is very similar to our own, namely that D.F.'s improved performance is based on access to dorsal-stream processing. The alternative, namely that motor-induced attention could have enhanced the representation in the ventral pathway, is highly unlikely given that recent MRI and fMRI evidence has shown that the relevant form areas of the ventral pathway are destroyed in patient D.F. (James et al., 2003).
The findings of this study not only show that the perceptual system can successfully access information from the dorsal stream, they also suggest that the intact grasping behaviour of D.F. cannot be taken as evidence for implicit processing of shape per se in the dorsal stream. In the first studies on D.F.'s grasping behaviour (Goodale et al., 1991, 1994b) her ability to adjust her grip to the shape of the target object might seem to suggest that she still had implicit access to shape information. In reality, however, Goodale et al. (1991) only reported that D.F. could use width information to guide her grasping movements. In traditional grasping studies the effects of shape information and width information are hard to disentangle because changes in shape are only likely to have an effect on grasp aperture when those changes are linked to changes in object width (Jeannerod, 1996; Smeets & Brenner, 1999; Rosenbaum et al., 2001). However, using the paradigm from Experiment 5, it was possible to disentangle the effects of shape and width. Combining the shape-discrimination task with the grasping task allowed us to assess the visual information that was present in the dorsal stream independently of its effect on the formation of the grasp aperture. We could therefore examine whether target objects of identical width but different shape would also improve shape-discrimination performance. We expected that if shape information is used during a grasping movement and thus represented in the dorsal stream, then improved shape-discrimination performance should be observed even for objects of different shape but identical width. However, we found that D.F.'s performance in this condition is not better than chance, suggesting that she is only processing width information in this grasping task, and not shape. This means that D.F.'s preserved grasping ability cannot be taken as evidence for the processing of shape information in the dorsal pathway. This conclusion is consistent with an earlier study showing that D.F.'s ability to choose optimal grip points is impaired in a condition where ‘true’ shape processing is required because no single dimension is sufficient to inform the best choice of grip points (Goodale et al., 1994a; Carey et al., 1996). However, it is worth noting that in non-human primates evidence for shape processing in the dorsal stream has been obtained (Sereno & Maunsell, 1998).
Although it seems likely that D.F.'s improved perceptual performance is based on information from the dorsal stream, it remains unclear as to exactly how this improvement could come about. If we assume that only the ventral stream provides visual information to our cognitive system then we also have to assume that it is via the modulation of activity in preserved parts of D.F.'s ventral stream that the dorsal representation exerts its effect on the perceptual judgement. Candidates for such preserved parts of D.F.'s ventral stream have been identified in a recent fMRI study (James et al., 2003). In that study, it was shown that if grey-scale or colour images of objects are used, both D.F.'s primary visual cortex and areas of the lingual and fusiform gyri become active, whereas these areas remain inactive when line-drawings are used. These regions of the ventral stream might be able to support the width discrimination required in our perceptual tasks if additional information from the dorsal stream is provided. Alternatively, it is quite possible that the cognitive system can gain direct access to the visual representation in the dorsal stream. In fact, functional imaging studies have shown that some of the more posterior areas in the dorsal stream (notably area cIPS) are activated during object recognition tasks (Faillenot et al., 1997, 1999; James et al., 2000, 2002). Unfortunately, on the basis of our findings it is not possible to decide whether it is through dorsal/ventral interaction or through direct access to this dorsal activity that the improvement of perceptual performance with concurrent visuomotor activity is achieved.
One notable aspect of our results is that D.F.'s discriminative performance was able to improve by virtue of visuomotor processing prior to the execution of her grasping response. This seems to suggest that the relevant representation of object attributes in the dorsal stream becomes available during the programming of the visuomotor act. This would be at odds with a characterization of the dorsal stream as a fast visual processing stream involved only in the online guidance of movements (Rossetti et al., 2003). However, there is evidence from patients (Perenin & Vighetto, 1988; Milner et al., 2003; Schindler et al., 2004) and healthy subjects (Culham et al., 2003; Frey et al., 2005; Medendorp et al., 2005; Prado et al., 2005; Sigman et al., 2005) that the dorsal-stream involvement in visuomotor control is by no means restricted to tasks that require online visual guidance; indeed, it is also apparent in tasks where all the relevant visual information is available prior to movement execution. In this case, the dorsal stream is presumably involved in extracting the visuomotor parameters used for the motor programme. A good example is provided by the visuomotor deficits of patients with lesions in the dorsal stream, namely patients with optic ataxia. These patients do have problems with the on-line visual guidance of movements (Pisella et al., 2000), but they also have problems at the movement programming stage in both their pointing behaviour (Milner et al., 2003) and in taking obstacles into account when reaching (Schindler et al., 2004). Thus, our assumption that the dorsal representation is already available during the stage of planning a movement is quite consistent with our current knowledge of the role of the dorsal stream in the control of movements.
Our finding that D.F.'s discriminative performance was already improved prior to the execution of the grasping response may also seem surprising in light of the results of the earlier study of D.F. by Murphy et al. (1996). They compared D.F.'s performance not only in a conventional shape-discrimination task but also in a visuomotor version where she had to indicate her choice by reaching for the correct target object. It was found that her shape-discrimination performance was superior in this visuomotor task. Murphy and colleagues argued, however, that D.F. had to rely on in-flight corrections to her reaching responses in order to achieve this superior discrimination performance. This reliance on in-flight corrections might seem to be inconsistent with our claim that the relevant information for the width discrimination becomes available prior to the execution of the reaching movement. We do not believe, however, that Murphy et al.'s findings contradict our claim of the availability of width information during the planning stage. Rather, we think that the specific nature of the task used by Murphy and colleagues might explain why their results differed from ours. Murphy et al. used a task where two objects were presented in any given trial, and thus D.F. first had to decide which of the two objects corresponded to the target shape (e.g. a square), and then move towards the selected target object. Thus, in this task D.F.'s initial movement programme would have been determined by her initial perceptual judgement, and therefore quite possibly dominated by degraded information from the ventral stream (such information was sufficient to result in a greater than chance discrimination in their task, as the authors used an easier pair of shapes than ours). Veridical dorsal-stream information about the target object may have become available later on and triggered the reported mid-flight corrections. This is in contrast to our study where no target selection was required. Thus, in our study D.F. could prepare the reach-to-grasp movement at the beginning of the trial, and the information processed during the motor preparation phase was then available to influence her verbal report. In our view, these differences between the tasks used in the two studies could explain why in contrast to Murphy et al. we found evidence of visuomotor facilitation already at the motor programming stage.
In conclusion, our findings suggest that the perceptual system may be able to gain access to an object's visuomotor processing in the dorsal stream, when more direct routes are unavailable due to brain damage. (It must be admitted, of course, that better ‘discrimination’ does not necessarily imply better ‘perception’, and instead D.F. could have been employing some indefinable ‘implicit’ cues of the kind that enable blindsight patients to perform above chance in discrimination tasks.) Our results also suggest that the adjustment of D.F.'s grip aperture to different types of rectangular objects is based on width rather than on shape information. On this interpretation, the finding of intact grip adjustment in D.F. would only reflect implicit size processing, rather than shape processing per se, in the human dorsal stream.
Acknowledgements
This work was supported by grants from the Leverholme Trust (Grant number: F/00128/O) and the UK Medical Research Council (Grant number G0000003).
Abbreviations
-
- LO
-
- lateral occipital area
-
- MRI
-
- magnetic resonance imaging