

Plant invasion is associated with higher plant–soil nutrient concentrations in nutrient-poor environments
Abstract
Plant invasion is an emerging driver of global change worldwide. We aimed to disentangle its impacts on plant–soil nutrient concentrations. We conducted a meta-analysis of 215 peer-reviewed articles and 1233 observations. Invasive plant species had globally higher N and P concentrations in photosynthetic tissues but not in foliar litter, in comparison with their native competitors. Invasive plants were also associated with higher soil C and N stocks and N, P, and K availabilities. The differences in N and P concentrations in photosynthetic tissues and in soil total C and N, soil N, P, and K availabilities between invasive and native species decreased when the environment was richer in nutrient resources. The results thus suggested higher nutrient resorption efficiencies in invasive than in native species in nutrient-poor environments. There were differences in soil total N concentrations but not in total P concentrations, indicating that the differences associated to invasive plants were related with biological processes, not with geochemical processes. The results suggest that invasiveness is not only a driver of changes in ecosystem species composition but that it is also associated with significant changes in plant–soil elemental composition and stoichiometry.
Introduction
The structure, diversity, and production capacity of terrestrial ecosystems are strongly linked to the concentrations and stoichiometric ratios in the different ecosystemic compartments and the soil availability of nitrogen, phosphorus, and potassium (Sterner & Elser, 2002; Reich & Oleksyn, 2004; Elser et al., 2007; Vitousek et al., 2010; Sardans et al., 2011; Peñuelas et al., 2013; Sardans & Peñuelas, 2015). Most drivers of global change, such as increasing atmospheric CO2 concentrations, N eutrophication, drought, warming, or land-use changes, change those elemental compositions and stoichiometries of ecosystemic compartments and those relationships with ecological processes and species composition (Seabloom et al., 2006; Elser et al., 2010; Sardans & Penuelas, 2012; Sardans et al., 2012; Peñuelas et al., 2013; Yuan & Chen, 2015). The growing success of invasive plants in many regions, 20% or more of plant species are exotics in many continental areas and 50% or more in islands (Seabloom et al., 2006); for example, plant invaders are affecting 405 000 km2 in United States (Seabloom et al., 2015), is an emerging driver of Global Changes; however, it has not received the same level of attention at this regard of the impacts on plant–soil nutrient concentrations (Hulme, 2009, 2015).
Previous studies have observed that several mechanisms involved in the uptake and nutrient-use efficiency by plants underlie the success of invasive plants (Daehler, 2003; González et al., 2010). The mechanisms seem to differ between nutrient-poor and nutrient-rich soils. In nutrient-poor soils, most studies suggest that the success of invasive plants depends on conservative strategies, such as a higher nutrient-use efficiency (Ostertag & Verville, 2002; Funk & Vitousek, 2007; González et al., 2010; Matzek, 2011), especially on short time scales (Funk & Vitousek, 2007), long nutrient residence times (Laungani & Knops, 2009), high resistance to low levels of nutrients (Kueffer, 2009; Schumacher et al., 2009), and high plasticity of stoichiometric ratios (González et al., 2010). In fact, all these traits are consistent with those expected in stress tolerant species (Grime, 1977), in this case by a stress due to nutrient limitation. The establishment of new symbiosis (Hiltbrunner et al., 2014) and the more effective use of existing symbiosis (Pringle et al., 2009) are other strategies frequently linked to plant invasiveness success, all them increasing the availability of limiting soil resources. In contrast, in nutrient-rich soils, there is an advantage of species with high rates of photosynthesis and growth (Schumacher et al., 2009; González et al., 2010), high reproductive outputs (González et al., 2010), large body size (Van Kleunen et al., 2010), low C : nutrient ratios in tissues (Peñuelas et al., 2009; Schumacher et al., 2009; González et al., 2010), low costs of foliar construction (Nagel & Griffin, 2001; González et al., 2010), large investments of N in photosynthetic production (Ehrenfeld, 2003; Shen et al., 2011), high capacities of nutrient uptake (Zabinski et al., 2002; Leffler et al., 2011; Peng et al., 2011), and high levels of plasticity in the acquisition of resources as a function of pulses in nutrient availability (Leffler et al., 2011). Nutrient uptake and all foliar traits enabling rapid rates of growth (Zabinski et al., 2002; Leishman et al., 2007) will thus help invading species to succeed when resources are not limited (Leishman et al., 2007; Peng et al., 2011). Some authors have claimed that, independently of growth conditions, invaders are more likely to have higher foliar areas, lower tissue construction costs, and greater phenotypical plasticity that increase the availability of soil resources (Daehler, 2003).
Invasive plant success has also been linked to differences in soil elemental composition. In a recent review, Pyšek et al. (2012) reported that 192 of 436 case studies on the effects of invasive plants on soil nutrient concentrations found higher concentrations, 72 found lower concentrations, and 158 found no significant differences. Sardans & Penuelas (2012), by analyzing 65 case studies, showed that most processes of invasion had higher availability of soil nutrients. In addition to these previous qualitative studies, Vilà et al. (2011) conducted a meta-analysis on the relationships of plant invasive success with soil condition showing that invasive success is related with higher soil C, N and P stocks.
There is, however, no general consensus on whether or not successful plant invaders have different elemental compositions than the native species, or, if present, whether differences are dependent or not on habitat nutrient richness. A quantitative study comparing plant, litter, and soil nutrient concentrations; that is, the whole plant–soil system, between invasive and their native competitors at the global scale, is missing. Moreover, there are no studies analyzing the differences for other important elements, such as K. In addition to the possible influence of soil nutrient richness, the possible influence of climate conditions on these relationships warrants investigation because climatic shifts affect invasive plant functional processes and in general invasion patterns (Lu et al., 2013; Zenni & Hoban, 2015) and thus could affect the differences in plant, litter, and soil nutrient concentrations between invasive and native plants at the global scale. In regions where climate evolves toward characteristics more favorable to plant production (higher MAP and/or MAT) and where invasive success is expected to be related to higher rates of nutrient uptake and in general to C and/or R ecological strategies (Grime, 1977), we should expect more investment of nutrients in plant growth and faster nutrient cycling rates in plant–soil system. Contrarily, in regions evolving toward more extreme and stressed climatic conditions, we should expect invasive success to be related to more conservative traits, less growth, traits typical of stress tolerator biological strategy (T strategy; Grime, 1977) that are less linked with higher-uptake capacity, but to a higher resorption and retention of nutrients in the system and consequently with higher nutrient concentrations in plant–soil system.
We have conducted a global meta-analysis of both the past and the most recent literature data on the nutrient concentrations in photosynthetic tissues, foliar litter, and soil with the aims to determine whether or not invasive plant success (i) is associated with different elemental compositions of photosynthetic tissues and foliar litter between successful invasive plants and their native competitors; (ii) is associated with changes in soil elemental composition and nutrient availability and stoichiometry; and (iii) how these associations, if exist, depend on soil nutrient concentrations and availabilities, and climatic condition.
Materials and methods
Data collection
We searched the ISI Web of Science using combinations of the following keywords: alien, availability, available, carbon, concentration, C : K, C : N, C : P, foliar, invasion, invasive, leaf, needle, nitrogen, N : K, N : P, phosphorus, plant, potassium, P : K, ratio, soil, solution, stoichiometric, stoichiometry, success. We only selected studies providing the same equivalent information for invasive successful species and their native competitors. Moreover, we only analyzed plant, litter, and soil variables with a minimum of 45 different reports that included the information for invasive and the respective native species. These variables finally included N and P concentration and C : N concentration ratio in photosynthetic tissues, foliar litter N concentration and C : N concentration ratio, and soil total C, N and P concentrations, total soil C : N concentration ratio, soil P-Olsen and soil extractable K+, 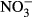 , and
, and 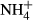 concentrations. In the few studies with different temporal data, we used the average mean values. Finally, only field nonmanipulative studies have been considered. Applying these criteria, we obtained 215 reports with 1233 observations across the world (Fig. S1).
concentrations. In the few studies with different temporal data, we used the average mean values. Finally, only field nonmanipulative studies have been considered. Applying these criteria, we obtained 215 reports with 1233 observations across the world (Fig. S1).
Climatic data
We extracted climatic data for each study site from the worldclim database (Hijmans et al., 2005). This database provides global maps of interpolated variables of climatological variables extrapolated from extensive climatic time series (from 1950 to 2000), with a spatial resolution of 30 arc seconds (~1 km at the equator). We used MAT and MAP as climatic predictor variables.
Data analyses
We examined the effects of invasive plant success on the differences of photosynthetic tissues elemental compositions and stoichiometries and soil nutrient status between successful invasive plants and their native competitors by calculating the ln response ratios from each study as described by Hedges et al. (1999). The natural ln response ratio (lnRR) was calculated as ln (Xi/Xn) = lnXi − lnXn, where Xi and Xn are the values of each observation in the invaded soil or invasive plant and in the corresponding native situation, respectively. The sampling variance for each lnRR was calculated as ln[(1/ni) × (Si/Xi)2 + (1/nn) × (Sn/Xn)2] using the r package metafor 1.9−2, where ni, nn, Si, Sn, Xi, and Xn are the invasive and native sample sizes, standard deviations, and mean response values, respectively. The natural ln response ratios were determined by specifying studies as random factors using the rma model in metafor. The effects on soil elemental variables and the difference between the elemental compositions of invasive and native plants were considered significant if the 95% confidence interval (CI) of lnRR did not overlap zero. All these statistical analyses were performed in r 3.1.2 (R Core Team, 2015). Despite most studied variables, there was a low proportion of studies containing N2-fixing species, and we performed these analyses twice, once with the entire data another one with and after the removal of the studies that contained N2-fixing plant species for detecting the possible importance of N2-fixing capacity in the ln response ratio effect of the plant and soil variables studied. We analyzed variables with more than 45 observations available at the global scale. The number of reports and observations used by statistical analyses of each studied soil, plant, and litter variable are shown in Figs 1 and 2, and described in Tables S1–S3.


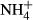 ), nitrate (
), nitrate ( ), P-Olsen, and total P, N, and C and the soil C : N ratio to plant invasion for the entire data set (including data for N2-fixing plants) (a) and excluding the data for N2-fixing plants (b). Values are means and 95% confidence intervals. Plus (+) and minus (−) signs represent positive and negative log response ratios, respectively, when the corresponding ln response ratios confidence intervals do not overlap with zero value. Zero in the X-axes represents neutral response ratio that means equal values in native than in invasive species. The numbers between brackets indicate the number of articles and studies, respectively, used in the meta-analysis of each variable.
), P-Olsen, and total P, N, and C and the soil C : N ratio to plant invasion for the entire data set (including data for N2-fixing plants) (a) and excluding the data for N2-fixing plants (b). Values are means and 95% confidence intervals. Plus (+) and minus (−) signs represent positive and negative log response ratios, respectively, when the corresponding ln response ratios confidence intervals do not overlap with zero value. Zero in the X-axes represents neutral response ratio that means equal values in native than in invasive species. The numbers between brackets indicate the number of articles and studies, respectively, used in the meta-analysis of each variable.We also examined whether the differences in the ln response ratio of plants and soils depend on environmental circumstances such as climate or soil total nutrient concentration and soil available nutrient concentration. For these analyses, we related the ln response ratio effect mentioned above (lnRR) with climatic variables at each study site. We used MAP and MAT data from the worldclim database (Hijmans et al., 2005). We also tested whether lnRR is dependent on native plant and soil total nutrient concentrations and soil nutrient availability. We conducted a regression of the ln response ratio of the soil N concentration (ln invaded soil N value – ln native soil N value) relative to the concentration in the natural (native site) soil (ln native soil N value). In the case of foliar plant tissues, nutrient concentration has generally been well correlated with soil nutrient availability across natural gradients or fertilization experiments (Porder et al., 2005; Alvarez-Clare & Mack, 2015). We thus used the native foliar concentrations as a proxy of site soil availability to relate the possible differences in the ln response ratio effect in foliar and foliar litter variables (ln invasive plant N value – ln native plant N value), with the corresponding variable availability in soil (ln native foliar N value). We used regression type II for these analyses, because both dependent and independent variables were interchangeable and random, so the error of the independent variable could not be neglected. We ran a standardized major axis method (SMA) using the smatr package (Warton et al., 2006) (http://www.bio.mq.edu.au/ecology/SMATR).
Finally, in the cases of total soil N concentration, soil P-Olsen and foliar N and P concentrations, for which we have the larger number of observations, we divided the observations of each one of these variables according with their values in native soils or plants in three groups with similar number of observations. Thus, the groups corresponded to low, intermediate, and high values in native conditions as a proxy of site nutrient richness. Thereafter, we conducted an one-way anova with Bonferroni post hoc test to detect possible differences in the ln response ratio among the three groups.
Results
Differences in photosynthetic tissues and foliar litter
A meta-analysis of the entire data set indicated that invasive plant species had higher N (z = 8.93, P < 0.0001) and P (z = 3.44, P < 0.001) concentrations (41% and 32%, respectively) and lower (26%) C : N ratios (z = −5.02, P < 0.0001) in their photosynthetic tissues than the native competitors (Fig. 1a). An analysis of the same data set but without excluding N2-fixing species also indicated higher N (z = 6.57, P < 0.0001) and P (z = 2.67, P < 0.01) concentrations (29% and 32%, respectively) and lower (22%) C : N ratios (z = −4.84, P < 0.0001) in the photosynthetic tissues of the invasive species (Fig. 1b). The N concentration and the C : N ratio in foliar litter were, however, not significantly different either for the entire data set (Fig. 1c) or when the data for the N2-fixing plant species were excluded (Fig. 1d). Not significant differences were either found for litter P concentrations (only 13 observations, data not shown).
Differences in soil conditions
The soil concentrations of extractable K (z = 2.53, P < 0.05), soluble nitrate (z = 7.40, P < 0.0001), P-Olsen (z = 2.83, P < 0.01), total N (z = 4.34, P < 0.0001), and C concentrations (z = 3.62, P < 0.001) were higher (13%, 117%, 21%, 19%, and 12%, respectively) in soils of invasive plants than in soils of their corresponding native competitor species. The concentration of soluble ammonium was also marginally (z = 1.81, P = 0.07) higher (11%) in the soils of the invasive than the native species. The ln response ratio effects on the soil C : N ratio and total P concentration were not statistically significant.
An analysis of the same data set but without the data for the N2-fixing species produced similar results (Fig. 2b). The soluble nitrate (z = 6.37, P < 0.0001), P-Olsen (z = 2.83, P < 0.001), total N (z = 2.32, P < 0.05), and C (z = 3.13, P < 0.001) concentrations were higher (118%, 27%, 10% and 7%, respectively) in the soils of the invasive plants than in the soils of the native competitors. The concentration of extractable K was marginally (z = 1.80, P = 0.072) higher (11%) in the soils of the invasive species.
Ln response ratios along gradients of nutrient availability and climate
The ln response ratio of total N and P concentrations in photosynthetic tissues of invasive plants were negatively correlated to the corresponding values for the photosynthetic tissues of the native plant competitors (Fig. 3a, b). The ln response ratio of foliar N concentration was positively different from zero in sites with low and intermediate values, whereas for foliar P concentrations the ln response ratio was only positively different from zero in sites with low values (Fig. S2a, b). No significant relationships were observed between foliar litter N and P ln response ratio and the corresponding values for the foliar litter of the native plant competitors (Fig. 3c, d).

The ln response ratio for soil total N, P-Olsen, soluble nitrate, and extractable K concentrations in invaded soils were negatively correlated with the corresponding values in the soils of the native plant competitors (Fig. 4a–d). For soil nitrate concentration, total N concentration and soil P-Olsen, the ln response ratio was positively different than zero in sites with low and intermediate values, whereas for soil available K+ the ln response ratio was positively different than zero only in sites with low values (Fig. S3).

 and the site soil
and the site soil 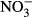 concentration (a), between the ln response ratio of soil total N and site soil total N concentration (b), between ln response ratio of soil P-Olsen and site soil P-Olsen concentration (c) and between ln response ratio of soil K+ concentration and site soil K+ concentration (d). Dotted line highlights the zero value of ln response ratio (equal values of the corresponding variable for native and in invasive species or for soils under them.
concentration (a), between the ln response ratio of soil total N and site soil total N concentration (b), between ln response ratio of soil P-Olsen and site soil P-Olsen concentration (c) and between ln response ratio of soil K+ concentration and site soil K+ concentration (d). Dotted line highlights the zero value of ln response ratio (equal values of the corresponding variable for native and in invasive species or for soils under them.Interestingly, few relationships between climatic gradients and ln response ratio effects were detected. MAT was positively but weakly correlated with the ln response ratios for soil total N concentration (R = 0.27, P < 0.001) and with N concentration in photosynthetic tissues (R = 0.16, P < 0.05). MAP was positively and also weakly correlated with the ln response ratio for soil soluble nitrate concentration (R = 0.25, P < 0.01) (Fig. S4).
Discussion
Our study showed higher N and P concentrations in the photosynthetic tissues of invasive species in nutrient-poor environments. These higher concentrations were found in photosynthetic tissues but not in foliar litter, suggesting a higher N and P resorption capacity in resource-poor than in resource-rich environments. These results are consistent with previous studies observing that the competitive advantage over native plant species competitors and the success of invasive plants in resource-poor environments has frequently been correlated with a more conservative use of nutrients, higher residence time due to higher nutrient resorption capacities (Ostertag & Verville, 2002), and higher photosynthetic nitrogen use efficiency (Ens et al., 2015).
The soils under the invasive plants had higher soil P-Olsen, soluble nitrate, and potassium concentrations and therefore higher availability of the three most important soil macronutrients for plant growth. The higher soil  concentrations in soils under invasive species than under their native competitors are consistent with previous studies observing a positive relationship between soil
concentrations in soils under invasive species than under their native competitors are consistent with previous studies observing a positive relationship between soil  concentration and the intensity of plant species invasive success (Gilliam, 2006). The studies compiled in this meta-analysis did not allow a clear determination of whether these higher concentrations were the cause or the effect of the success of invasive plant species. The studies that have experimentally tested whether soil differences were the cause or the consequence of plant invasion, however, have reported that soil differences were mainly due to the effect of the success of the invasive species (Li et al., 2006; Dassonville et al., 2008; Elgersma et al., 2011; Lee et al., 2012; Kuebbing et al., 2014; Stark & Norton, 2014). A few number of reports that have studied the changes in soil conditions during 4 (Belnap et al., 2005) and 7 (Hawkes et al., 2005) years have observed that the invasive species changed soil conditions over time. Several studies have also observed a direct impact of invasive plant establishment on soil function such as increases in soil enzymatic activities associated with increases in some soil elemental concentrations (Hawkes et al., 2005; Allison et al., 2006; Caldwell, 2006; Aragón et al., 2014; Kuebbing et al., 2014), mineralization (Haubensak & Parker, 2004; Fickbohm & Zhu, 2006; Li et al., 2006), and respiration (Souza-Alonso et al., 2015). Other studies, although fewer than the above, did not observe these differences in soil enzymatic activity and mineralization (Zabinski et al., 2002; Meisner et al., 2011) or found different results depending on species and site (Koutika et al., 2007) or on the enzymatic activities (Chacón et al., 2009). Kulmatiski et al. (2006) in 660 experimental plots in abandoned croplands (from 50 to 7 years ago) with different management histories observed that invasive success explained the soil C, N, and P concentrations more significantly than the previous agricultural histories, suggesting that the invasive plants facilitated their own growth by maintaining beneficial fungal communities and fast nutrient cycling rates.
concentration and the intensity of plant species invasive success (Gilliam, 2006). The studies compiled in this meta-analysis did not allow a clear determination of whether these higher concentrations were the cause or the effect of the success of invasive plant species. The studies that have experimentally tested whether soil differences were the cause or the consequence of plant invasion, however, have reported that soil differences were mainly due to the effect of the success of the invasive species (Li et al., 2006; Dassonville et al., 2008; Elgersma et al., 2011; Lee et al., 2012; Kuebbing et al., 2014; Stark & Norton, 2014). A few number of reports that have studied the changes in soil conditions during 4 (Belnap et al., 2005) and 7 (Hawkes et al., 2005) years have observed that the invasive species changed soil conditions over time. Several studies have also observed a direct impact of invasive plant establishment on soil function such as increases in soil enzymatic activities associated with increases in some soil elemental concentrations (Hawkes et al., 2005; Allison et al., 2006; Caldwell, 2006; Aragón et al., 2014; Kuebbing et al., 2014), mineralization (Haubensak & Parker, 2004; Fickbohm & Zhu, 2006; Li et al., 2006), and respiration (Souza-Alonso et al., 2015). Other studies, although fewer than the above, did not observe these differences in soil enzymatic activity and mineralization (Zabinski et al., 2002; Meisner et al., 2011) or found different results depending on species and site (Koutika et al., 2007) or on the enzymatic activities (Chacón et al., 2009). Kulmatiski et al. (2006) in 660 experimental plots in abandoned croplands (from 50 to 7 years ago) with different management histories observed that invasive success explained the soil C, N, and P concentrations more significantly than the previous agricultural histories, suggesting that the invasive plants facilitated their own growth by maintaining beneficial fungal communities and fast nutrient cycling rates.
Our results showed a general globally higher soil total N concentration under invasive plants than under their native competitors but we did not observe a higher total P concentration. Cycling and concentrations of soil N mainly depend on biological processes, whereas mineral rocks are the sources of soil P, and its soil total concentration is primarily driven by physicogeochemical processes (Gómez-Aparicio & Canham, 2008; Vitousek et al., 2010; Peñuelas et al., 2013). Both N and P are important soil components that could be involved in facilitating plant invasion, but only soil total N concentration can thus be associated mostly with the biological process of plant invasion. This fact is consistent with the hypothesis that the differences between the soils under invasive and native plants are most likely due to the effects of species invasion itself. Rather surprisingly, the effects of plant invasions on soil and plant N concentrations, C : N ratios, and most other significant stoichiometry parameters were not different when including N2-fixing plants than when excluding them from the global analysis. Changes in soil physical conditions or in microbial communities, including soil N-fixing microbes, could be involved, warranting further research.
The differences in soil total C and N, and in N, P, and K availabilities and in N and P concentrations in photosynthetic tissues between invasive and native species decreased with increasing values of the corresponding variables in natural native conditions to the point that the differences disappeared in resource-rich environments. These lower differences in resource-rich environments could be due to the higher nutrient uptake in invasive species being counteracted by its higher growth capacity and the corresponding dilution effect. In nutrient-rich sites, moreover, native species are also highly competitive, having traits that enable native plants to be very effective in taking up resources. On the other hand, the frequently observed higher mineralization capacity and enzyme activity under invasive than under native species in nutrient-rich soils (Allison et al., 2006; Gómez-Aparicio & Canham, 2008; Aragón et al., 2014) would increase the rates of nutrient released from organic matter, but this would be also counteracted by the higher plant nutrient uptake so that soil nutrient concentrations would remain similar than under native species.
Climate variables had few correlations with the studied ln response ratio of the studied variables. MAT had weak but positive correlation with ln response ratio of total soil N concentration and N concentration in plant tissues and MAP had also a positive relationship with soil nitrate ln response ratio. These results suggest thus that climatic conditions are less influential on the ln response ratio of the studied soil total and available nutrient concentrations than the environmental nutrient richness.
Summarizing, this is the first study that has analyzed globally the association between plant invasion and nutrient concentration and stoichiometry of photosynthetic tissues, leaf litter, and soils. Invasive plant species had globally higher N and P concentrations in photosynthetic tissues but not in foliar litter, in comparison with their native competitors. Invasive plants were also associated with higher soil C and N stocks and N, P, and K availabilities. The differences in N and P concentrations in photosynthetic tissues and in soil total C and N, soil N, P, and K availabilities between invasive and native species decreased when the environment was richer in nutrient resources. These global trends may be explained by (i) larger differences in resorption and nutrient-use efficiency between invasive and native species in nutrient-poor environments, and (ii) a higher competitive capacity associated with larger nutrient uptake and plant growth capacity with a dilution effect in invasive than in native species when environments become richer in resources. Moreover, some other mechanisms such as enhancement of soil enzymatic activity and mineralization, and more effective symbiotic relationships can be also involved in these global trends. Clearly determining whether invasive plant success is the cause or the consequence of soil elemental composition and nutrient availability is currently not possible, but research up to now suggests that these plant and soil nutritional changes are more the consequence than the cause of plant invasion. Plant invasiveness should thus not be neglected as a driver of global change in plant–soil elemental and stoichiometric composition and soil fertility.
Acknowledgements
The authors would like to acknowledge the financial support from the European Research Council Synergy grant ERC-SyG-2013-610028 IMBALANCE-P, the Spanish Government grant CGL2013-48074-P, and the Catalan Government grant SGR 2014-274.
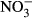 concentration (a), soil total N content (b), soil P-Olsen (c) and and soil K+ concentration (d) at different levels of the variable in the native environment.
concentration (a), soil total N content (b), soil P-Olsen (c) and and soil K+ concentration (d) at different levels of the variable in the native environment.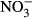 concentration with ln MAP (c).
concentration with ln MAP (c).
