


Flow intermittency decreases nestedness and specialisation of diatom communities in Mediterranean rivers
Summary
- Due to their ubiquity and highly efficient passive dispersal, benthic diatoms are an ideal group in which to explore the drivers of community spatial structure and the ultimate links between such patterns and species autoecology. However, studies addressing these issues have largely overlooked unicellular taxa to date.
- We examined the spatial order (i.e. nestedness) of a diatom meta-community of 122 lotic sites in the north-east Iberian Peninsula and the relationship between the contributions of individual species to nestedness and their niche breadths [measured using outlying mean index (OMI) analyses].
- We observed a significantly nested pattern in the meta-community (T = 14.05, NODF = 22.57), although idiosyncratic species accounted for a substantial fraction (34%) of the global species pool. Hydrological stability was identified as the main driver of nestedness. Highly stable, moderately stable and intermittent sites differed in nestedness and in alpha diversity, with intermittent sites being the most species poor and the least nested.
- We also observed a significant association between nestedness and niche attributes, with idiosyncratic species exhibiting wider distributions and larger niche breadths compared with nested species. Overall, our results indicate that diatom communities inhabiting hydrologically stable rivers present a higher level of order in community spatial pattern and a higher proportion of specialist taxa than communities in intermittent streams.
- Because recent hydroclimatic models predict an increase in stream intermittency in the Mediterranean region, our findings may help anticipate future diatom biodiversity patterns resulting from global change. We suggest that the current need to protect a diverse network of sites (as opposed to a few species-rich sites) will be even greater in the near future because local communities are not only probably to become taxonomically and functionally poorer but also less predictable, making it more difficult to implement efficient programmes for biodiversity conservation at the regional scale.
Introduction
Since Diamond's (1975) seminal work on community assembly rules, much research in community ecology has focused on the alternate paradigms that govern patterns of community assemblage. Within this context, both the ‘island paradigm’ (i.e. assembly rules are governed by dispersal-driven mechanisms) and the ‘trait-environment paradigm’ (i.e. niche assembly rules are the major force) have coexisted to date (Weiher & Keddy, 2001). Whereas the former paradigm focuses on processes such as immigration and extinction, the latter emphasises environmental filters and species autoecology. However, as noted by Soininen (2008), the two conceptual frameworks are not mutually exclusive. Because traits of individual species determine the ability to colonise new habitats or the probability of extinction, the autoecology of a particular species may be related to the spatial structure it exhibits (Mcabendroth et al., 2005).
When comparing communities from different sites, many matrix configurations are possible (Almeida-Neto et al., 2008). However, all situations in which communities differ can be described by a combination of two mechanisms, namely species replacement (i.e. turnover) and species gain or loss (i.e. nestedness) (Baselga, 2010). Therefore, nestedness analyses measure the degree of order in species gain and loss. Nested structures occur when assemblages of species-poor sites are subsets of the assemblages of species-rich sites. In this way, nestedness quantifies the overlap in species composition between high and low-diversity sites (Atmar & Patterson,1993). Nestedness is a feature of the meta-community level, being seen among a set of local communities that are linked by dispersal of multiple potentially interacting species (Leibold et al., 2004). This pattern has been observed in terrestrial and aquatic environments for many taxonomic groups, from plants to vertebrates (Patterson & Atmar, 1986; Wright et al., 1998; Sasaki et al., 2012; Ruhí et al., 2013).
Several mechanisms have been proposed to elicit nested patterns in meta-communities, such as selective extinction, passive sampling, selective colonisation, nested habitats and selective environmental tolerances (Ulrich, Almeida-Neto & Gotelli, 2008). Beyond the processes responsible for nestedness, identifying these patterns may have important implications for biodiversity conservation. A meta-community displaying a highly nested structure would allow conservation programmes to focus on the protection of a few rich habitats because those sites would contain the full set (or source pool) of species. However, strong nestedness is not the rule in nature; indeed, unexpected presences and absences of species are commonly observed, regardless of the taxonomical group (e.g. Soininen, 2008; Karthick, Mahesh & Ramachandra, 2011; Ruhí et al., 2013). Idiosyncratic species are those departing from the nested pattern, appearing unexpectedly in species-poor sites and/or being absent from species-rich sites (Atmar & Patterson, 1993). Within this context, recent studies have indicated that even the most depauperate sites sometimes contain taxa not found elsewhere within a region (Sasaki et al., 2012).
In contrast to what has been observed in larger taxa (e.g. plants and vertebrates), the macroecological patterns of unicellular organisms have been largely overlooked (but see Heino & Soininen, 2005, 2007; Soininen, 2008). However, an increasing body of research supports the idea that as with macroorganisms, microorganisms also exhibit biogeographical patterns with no strict evidence of ubiquitous global distribution (Hillebrand et al., 2001; Heino & Soininen, 2005). In the case of diatoms, historical processes (i.e. colonisation, extinction, dispersion, migration) may have shaped their global diversity patterns (Vyverman et al., 2007). Although there is no evidence for a ubiquitous global distribution, freshwater diatoms show relatively efficient passive dispersal at the within-region scale (Soininen & Heino, 2007). High between-patch dispersal has been proposed to reduce nested patterns (e.g. in aquatic invertebrates, Boecklen, 1997; Wright et al., 1998), due to a homogenisation of assemblage composition. Thus, it is expected that efficient dispersal will decrease the pattern of nestedness at this scale.
Diatoms are a siliceous class of unicellular algae known to be very sensitive to chemical conditions. They are distributed throughout the world in nearly all types of aquatic systems and usually account for the highest number of species among the primary producers in aquatic systems (Pan et al., 1999). Different diatom taxa exhibit different optima and tolerances to environmental factors, and their distributional patterns respond to a multitude of different factors, ranging from biogeographical to biogeochemical and human influenced (Potapova & Charles, 2002). Moreover, the structural elements in their siliceous cell walls allow for reliable taxonomic determination at specific and subspecific levels (Sabater & Admiraal, 2005). Thus, diatoms are an ideal model group in which to explore the drivers of spatial patterns of passively dispersed microorganisms, as well as to identify the ultimate links between individual species' contributions to such patterns.
The composition and abundance of algal communities are affected by multiple interacting factors, such as water chemistry, light availability, variations in temperature, water velocity, substrata type and grazing. These factors vary spatially and temporally, and amongst them, water flow is the physical factor affecting most other physicochemical features (Stevenson, 1996). For instance, Mediterranean streams are characterised by periods of hydrological stability that are interrupted by floods or droughts (Sabater et al., 2008). Seasonality and variability in rainfall is the principal attribute of the Mediterranean-type climate (Gasith & Resh, 1999), which causes low-flow periods during late spring and summer, and higher flow periods in late autumn and winter (Butturini et al., 2003). As a result, headwater and middle-order streams often become intermittent or have intermittent reaches (Lake, 2003), or have areas of reaches that are dry for extended periods (Sabater & Tockner, 2010). Intermittency influences the survival and performance of stream biota (Boulton, 2003; Datry, Arscott & Sabater, 2011) and controls the effects of small-scale heterogeneity (Tornés & Sabater, 2010). Specifically, droughts are known to have a profound impact on the densities and size- or age-structure of populations, on community composition and diversity, and on ecosystem processes (Lake, 2003). For instance, it has been suggested that very few diatom species are abundant and dominant in dry riverbeds, with these remaining species being highly tolerant (Mathooko et al., 2005). Disturbances caused by local flow fluctuations confer heterogeneity to the structure of benthic algae, with successional age being inversely related to disturbance frequency (Ledger et al., 2008). Moreover, hydrological disturbance may also provoke increases in beta diversity through differences among communities in disconnected habitat patches (Larned et al., 2010). As nestedness has been related to assemblage stability (Atmar & Patterson, 1993), this pattern is expected to increase with hydrological stability (Florencio et al., 2011). Extremely low-flow periods are probably to increase in the Mediterranean basin due to the influence of current climate change (Giorgi & Lionello, 2008) and human-related disturbances (Lehner et al., 2006). As a result, the running fresh waters of the Mediterranean and other semiarid regions are probably to experience a loss of habitats and associated biodiversity (Boix et al., 2010; Timoner et al., 2012).
The overarching aim of this study was to explore nestedness of stream microorganisms, using benthic diatoms as a model group. Specifically, we were interested in identifying the extent to which environmental variables determine nestedness and which autoecological characteristics typify nested and idiosyncratic taxa. We hypothesised that (i) stream diatom communities will exhibit a significantly nested pattern, as has been observed in multicellular freshwater organisms; (ii) this ordered pattern will increase with hydrological stability because the presence and absence of unexpected species should be more frequent in disturbed systems than in more stable ones; (iii) as a consequence, a higher species dissimilarity (i.e. beta diversity) will be observed more often in idiosyncratic sites than in nested ones, regardless of differences in species richness (alpha diversity); and (iv) species greatly contributing to the nested structure will be characterised by narrow ecological tolerances, and idiosyncratic species will be generalists, as the latter should present a wider range of strategies conferring resilience from, and resistance to, hydrological disturbance.
Methods
Study area
The study area was composed of 122 stream and river sites in the north-east Iberian Peninsula (Fig. 1). We selected these sites because they cover long environmental gradients (Appendix S1). Ninety of the 122 sites coincided with those from the control network of the Catalan Water Agency (ACA), whereas 32 were added to have a better representation of reference sites. The study region is characterised by a large heterogeneity in geomorphology and altitudinal range. The sites included in the study ranged from siliceous high-mountain fluvial systems to coastal streams, comprising both calcareous and siliceous fluvial systems. Most of the basins considered were strongly influenced by the Mediterranean climate, with marked seasons and high interannual variability in rainfall (Gasith & Resh, 1999). Some of the streams and rivers show summer flow interruption. A detailed description of sites is available in Tornés et al. (2007). The spatial scale of the study is 31 896 km2, not too large to limit stream diatoms dispersal (see Soininen & Heino, 2007). Moreover, there are no relevant physical barriers impeding efficient passive dispersal of diatom species in the study area.

Sampling and species identification
We conducted the sampling surveys during the summer (July–August) of 2002 and spring (May–June) of 2003. Sampling sites were visited twice to include both high-flow (spring) and low-flow conditions (summer). Sampling and counting of diatoms followed European Standards (EN 13946, 2003; EN 14407, 2004). We randomly collected at least five stones from the stream bottom in riffle sections, and diatoms were scraped off the stones to a final area of 2–10 cm2. We preserved algal samples in 4% formaldehyde until analysis.
The environmental variables analysed included both the physicochemical characteristics of the water and habitat descriptors. We determined water temperature, conductivity and pH in situ, by means of WTW meters (Wissenschaftlich-Technische Werkstätten, Weilheim, Germany). We made qualitative observations of water transparency, water velocity and canopy cover. Chemical data analyses (nitrate and phosphate) were determined by molecular absorption spectrophotometry (Standard Methods 4110 and EPA Methods A-100). We used the channel width in each site, as well as the altitude, as basic physiographic descriptors. We derived altitude from the Digital Elevation Model (DEM, 30× 30 m) of the Institute of Catalan Cartography (ICC, www.icc.cat) using ESRI ArcGIS®. We assigned the hydrological stability of each site to one of three possible categories (i.e. highly stable, moderately stable or intermittent) following the classification made by the water authority (ACA, 2005, 2008). Highly stable sites are related to humid Mediterranean climate. These are large rivers with regular lateral water inflows. Moderately stable sites are also related to humid Mediterranean climate or to headwaters with high rainfall. They are permanent streams that rarely dry out. Intermittent sites are related to dry Mediterranean climate. Rainfall is scarce in these streams and they often dry out in summer or, in some cases, during most of the year.
We cleaned diatom frustules from the organic material using sulphuric acid, dichromate potassium and hydrogen peroxide. We mounted frustules on permanent slides using Naphrax (r.i. 1.74; Brunel Microscopes Ltd., Chippenham, Wiltshire, U.K.). We counted up to 400 valves on each slide by performing random transects under light microscopy using Nomarski differential interference contrast optics at a magnification of 1000 ×. We identified taxa to the species level according to Krammer & Lange-Bertalot (1991a,b; 1997a,b) and Lange-Bertalot (2001).
Data analyses
We constructed two data matrices. One was a presence–absence biotic matrix based on the detection of each species on either of the two visits; therefore, if a species was present on one or on both visits to a given site, it was considered to be present; otherwise, it was considered absent. This matrix consisted of 419 diatom species and 122 sites. Second, we averaged the values of each environmental variable from the two available values (i.e. the two visits for each of the sites) and used these data to construct an environmental matrix. The final environmental matrix consisted of 11 environmental variables per 122 sites. Environmental data (except pH and data expressed as categorical variables) were log10(X + 1)-transformed before analyses to reduce skewed distributions.
We first aimed to determine the overall nestedness of the biota matrix. The degree of nestedness of a given binary matrix can be computed using several metrics. We used the software ANINHADO, which orders matrices according to the number of presences per row and draws a diagonal between species presence and absence (Guimarães & Guimarães, 2006). ANINHADO computes two nestedness metrics: the classical matrix temperature (T; Atmar & Patterson, 1993) and the more recently proposed nestedness metric based on overlap and decreasing fill (NODF; Almeida-Neto et al., 2008). Whereas the popular T metric has the advantage of allowing for comparison between the observed nestedness of matrices, species and sites with values from published data, the NODF is recommended for its theoretical consistency and statistical behaviour (i.e. the metric is independent of the matrix size and shape). Therefore, in this study, we use both measures. T can be interpreted as a thermodynamic measure of disorder, ranging from 0° in ordered systems (maximum nestedness) to 100° in disordered systems. NODF varies from 0 (minimum nestedness) to 100 (maximum nestedness). Subsequently, we compared the observed nestedness (Nobs) with the predicted nestedness (Npre) made by four individual null models (with 1000 matrix randomisations each). The four null models were as follows: (i) presences assigned at random across the matrix; (ii) a fixed–fixed null model, with both column (species) sums and row (sites) sums fixed; (iii) only column sums fixed; and (iv) only row sums fixed. In the data analysis, if significant nestedness was observed with the fixed–fixed model, then it was also detected using the other three null models. Therefore, we selected the fixed–fixed model due to its lower incidence of type I errors (Almeida-Neto et al., 2008). We then determined whether individual sites and taxa were nested or idiosyncratic, using T. We considered idiosyncratic species or sites as those with higher T values than the original matrix values, and nested ones as those with T values lower than or equal to the original matrix values (Atmar & Patterson, 1993).
Second, we explored which environmental variables were positively related to nestedness (i.e. those eliciting low T values). To this end, we performed linear regressions between each quantitative environmental variable (nitrate, phosphate, water temperature, conductivity, pH, channel width and altitude) and the T values obtained in the former analyses. Relationships between qualitative measurements (water transparency, water velocity, canopy cover, hydrological stability) and nestedness (temperature T) were analysed by means of a generalised linear model (GLM), with temperature T as response variable and each environmental variable as predictor.
Third, we aimed to characterise nested and idiosyncratic sites with regards to their community attributes. Diatom species richness (i.e. alpha diversity) was calculated by summing all recorded species per site. The diatom species dissimilarity (i.e. beta diversity) was calculated using Sorensen index and PERMDISP, a multivariate test of homogeneity of dispersion that provides the distance of the composition of each site to the centroid. Sites with higher distances to the centroid have greater species dissimilarity with the rest of the sites (i.e. share less species) than sites presenting lower distances to the centroid. This method has been recently used to assess beta diversity patterns (Al-Shami et al., 2013). Differences in alpha and beta diversity between nested and idiosyncratic sites were tested with an analysis of variance (ANOVA), with alpha or beta diversity as dependent variables and nested structure (nested versus idiosyncratic sites) as fixed factor.
Finally, we aimed to address whether the magnitude of each species' contribution to nestedness was related or not to its autoecology. To this end, we calculated occurrence and niche breadth values for each taxon. Species occurrence was calculated as the proportion of sites where each species occurred using the biota matrix. Moreover, we combined the biota and the environmental matrices to calculate species niche breadths. The niche breadth for each taxon was obtained using outlying mean index (OMI) analysis (Dolédec, Chessel & Gimaret-Carpentier, 2000). We selected the ‘species tolerance’ (Tol) parameter, which measures the dispersion from the mean of the environmental variables used by a species, and hence represents its niche breadth. Within this context, species exhibiting a wide niche breadth tolerate a wider range of environmental conditions (i.e. they can be regarded as generalist species) than those presenting a narrow niche breadth (i.e. specialist species). This method has been widely employed in similar studies dealing with different taxonomical groups, from algae to invertebrates (e.g. Azeria & Kolasa, 2008; Soininen, 2008). Niche breadth was calculated for all species, and Monte Carlo permutation tests (999 permutations) allowed the statistical significance of the measure to be evaluated for each species. These analyses were performed with the functions ‘pcaivortho’, ‘niche’ and ‘rtest’ in the ade4 package (Dray & Dufour, 2007), written in R language. Finally, we examined differences in niche attributes between nested and idiosyncratic species using an ANOVA, with species tolerance as dependent variable and nested structure (nested versus idiosyncratic species) as fixed factor.
Nestedness was obtained with ANINHADO 3.0 (see www.guimaraes.bio.br/soft.html, Guimarães & Guimarães, 2006). Regression analyses, GLM and ANOVA were calculated with the software package PASW version 17 (SPSS, Chicago, IL, U.S.A). PERMDISP was run with PRIMER-E version 6.1.11 and PERMANOVA + version 1.0.1 (PRIMER-E Ltd., Ivybridge, U.K.). Outlying mean index (OMI) analyses and Monte Carlo permutations were performed with R software version 2.15.0 (R Foundation for Statistical Computing, Vienna, Austria).
Results
The diatom communities in the northeast Iberian Peninsula showed a significantly nested pattern. The observed temperature T (Tobs = 14.05) was significantly lower than that predicted by the matrix randomisations under the ‘fixed–fixed’ null model (Tpre = 42.41), indicating that the meta-community was highly nested. The same pattern of a more highly nested structure was observed using the NODF metric, which showed an observed global value (NODFobs = 22.57) that was much higher than that predicted by the null model (NODFpre = 11.86). Therefore, in both cases, the null hypothesis of no significant nested structure was rejected (P < 0.001). Nevertheless, a high number of idiosyncratic taxa were identified (143), accounting for the 34% of global species pool (419) (Fig. 2).

Hydrological stability and channel width were the environmental variables that were related to the degree of nestedness of the studied sites (Tables 1 & 2). Nestedness significantly differed among hydrological types (Fig. 3; F2,119 = 3.770, P = 0.026). The Games–Howell nonparametric post hoc test determined that significant differences occurred between highly stable and intermittent sites (P = 0.006), but there were no differences between highly and moderately stable sites (P = 0.079), or between moderately stable and intermittent sites (P = 0.187). The highly stable typology presented the highest proportion of nested sites, whereas the intermittent typology had the highest proportion of idiosyncratic sites. The gradient of hydrological stability was associated with that of increasing nestedness (i.e. increasing proportion of nested and decreasing proportion idiosyncratic sites), as we had predicted. Moreover, taxon richness differed among hydrological stability typologies. The mean [± standard error (SE)] of species richness in highly stable sites was 56.4 (±3.3), whereas it was 47.0 (±1.6) in moderately stable sites and 42.2 (±2.8) in intermittent sites. Hence, the intermittent sites, which had the highest proportion of idiosyncratic taxa, were also the poorest in terms of species richness.
| r2 | P | Coefficient | Standardised coefficient | |
|---|---|---|---|---|
| NO3−-N | 0.010 | 0.279 | −4.617 | −0.099 |
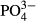 -P -P |
0.027 | 0.068 | 3.396 | 0.166 |
| pH | 0.001 | 0.691 | 1.460 | 0.036 |
| Conductivity | 0.005 | 0.447 | 2.145 | 0.069 |
| Water temperature | 0.000 | 0.934 | 1.153 | 0.008 |
| Width | 0.056 | 0.009 | −6.931 | −0.237 |
| Altitude | 0.004 | 0.511 | 1.236 | 0.060 |
| AIC | P | |
|---|---|---|
| Canopy cover | 980.28 | 0.455 |
| Water velocity | 979.95 | 0.552 |
| Water transparency | 978.08 | 0.427 |
| Hydrological stability | 970.37 | 0.021 |
- AIC, Akaike information criterion.

Differences existed between nested versus idiosyncratic sites in terms of species richness (i.e. alpha diversity) and spatial species dissimilarity (i.e. beta diversity). As predicted, idiosyncratic sites showed lower alpha diversity than nested sites (Fig. 4a; F1,120 = 95.168, P < 0.0001), whereas the pattern for beta diversity was the opposite, with idiosyncratic sites exhibiting higher values than nested sites (Fig. 4b; F1,120 = 36.343, P < 0.0001). The spatial dissimilarity of community composition occurred at higher rates among idiosyncratic than among nested sites, although idiosyncratic sites were poorer in terms of species richness than the nested sites.

The taxa that contributed most to the nested structure had low total occurrences in general, whereas idiosyncratic taxa tended to be present more frequently and across all hydrological typologies (Fig. 5). The association between nested structure (nested versus idiosyncratic species) and occurrence was significant (F1,417 = 128.168, P < 0.0001; see Appendix S2 for the complete table of occurrences and nestedness values). The most nested taxa (T < 0.01) were Achnanthes brevipes var. intermedia, Achnanthidium coarctatum, Halamphora holsatica, Gomphonema exiguum var. minutissimum, Melosira moniliformis, Fallacia cryptolyra, Fallacia forcipata and Navicula salinicola. These taxa were exclusively found in highly stable sites. At the other extreme, the most idiosyncratic taxa (T > 60) were Sellaphora seminulum, Fistulifera saprophila, Halamphora veneta, Nitzschia frustulum, Nitzschia desertorum, Nitzschia aurariae and Hannaea arcus. Most of these occurred in all three hydrological stability categories. Also, a small group of taxa with very high occurrences (>75%) was nested taxa. This group included Nitzschia palea, Navicula cryptotenella, Nitzschia dissipata, Cocconeis placentula, Amphora pediculus and Achnanthidium minutissimum.

The OMI analyses identified 147 from 419 taxa with significant niche breadth values (Monte Carlo permutations P < 0.05). Mean species niche breadth (±SE) of the whole biota matrix was 1.44 (±0.08), with values ranging from <0.1 (15 species) to >3.0 (four species) (see Appendix S2 for the complete table of niche breadth and nestedness values). When species were classified as nested or idiosyncratic, clear differences emerged in their mean niche breadths, with idiosyncratic taxa showing significantly larger niche breadths than nested taxa (Fig. 6; F1,145 = 22.660, P < 0.0001). Therefore, idiosyncratic taxa, which are widely distributed and most frequently associated with intermittent streams, were also more generalist than those found in highly stable sites.

Discussion
Causes of nestedness in benthic diatom meta-communities
Freshwater biota tends to present, in general, a high degree of nestedness (Heino, 2011). However, nestedness patterns across species and trophic levels in fresh waters have recently been described as showing a low congruence (i.e. low cross-taxon concordance) (Soininen & Köngäs, 2012). The diatom meta-community analysed in this study showed a relatively high proportion of species classified as idiosyncratic, in spite of its significant nested structure. The observed temperature T in this study (Tobs = 14.05) was relatively low, in line with, or relatively lower than the previous studies on diatom communities (T = 16.41 in Karthick et al., 2011; T = 28.4 in Soininen, 2008). These values are considerably higher, however, than other studies assessing nestedness in freshwater invertebrates. For instance, a T of 5.3 was obtained in a wetland macroinvertebrate meta-analysis comprising 447 wetlands in several ecoregions across the world (Batzer & Ruhí, 2013), and values as low as 8.9 were described from clam and snail assemblages in littoral areas of boreal lakes (Heino & Muotka, 2005). The passive dispersal of unicellular organisms could be associated with a lower, yet significant, nestedness than multicellular organisms with other dispersal strategies.
The differential colonisation of active and passive dispersers has been proposed as a major cause of nestedness in freshwater fauna. For instance, high species overlaps between poor and rich systems have been observed when comparing recent man-made wetland faunal communities with those inhabiting nearby natural wetlands (Ruhí et al., 2013). In those cases, natural systems act as a source of species for man-made systems because the latter would host subsets of the potential pool of recruiters. As a result, the variability in dispersal ability of colonisers would ‘naturally’ provoke a nested structure in those meta-communities. In contrast, the regional ubiquity and lack of differential colonisation of unicellular organisms (Heino & Soininen, 2005; Soininen & Heino, 2007) probably contribute to less ordered species gains and losses, and taxa may be able to appear virtually anywhere. Thus, although significant nestedness can occur in aquatic biota even when they are dominated by species with high interlocality dispersion rates (Rundle et al., 2002), it seems logical that highly efficient passive dispersal at the within-region scale (as have diatoms, Soininen & Heino, 2007) should reduce any type of non-random pattern.
The nestedness of local communities was significantly associated with hydrological stability (measured in three broad categories) and channel width. This is not unexpected as large systems are often those with relatively higher hydrological stabilities. Marked hydrological variability and flow interruption are characteristic of Mediterranean streams. Because this is a governing factor that summarises habitat availability, the chemical and physical environment, as well as the degree of associated grazing pressure (Sabater & Tockner, 2010), it is not surprising that the hydrological categories differed greatly in the number of idiosyncratic and nested species they hosted. An analysis of benthic diatom communities in 105 Finnish streams revealed only weak, non-significant relationships, between nestedness and environmental factors (Soininen, 2008). In contrast, habitat suitability has been described as one of the predominant determinants of nestedness in lentic systems (e.g. Heino & Muotka, 2005; Mcabendroth et al., 2005). Because Mediterranean rivers approach lentic conditions during low water flow and flow interruption (Sabater, 2008), we suggest that the relationship between nestedness and environmental characteristics observed in this study is at least partly due to this ‘lentification’. Our hypothesis that the presences and absences of ‘unexpected’ species (i.e. species disrupting the ordered pattern and thus contributing to idiosyncrasy) would be more frequent in systems prone to major abiotic disturbance (e.g. episodes of drying-up in Mediterranean streams) than in highly and moderately stable sites was therefore confirmed. The observed pattern is supported by the mechanisms of benthic algal extinction-colonisation dynamics, which are dependent on natural physical disturbance (Peterson, 1996).
It is important to note that in this study, the taxon list for each site encompassed two samplings, under both high- and low-flow conditions. We therefore propose that along with high- and low-flow conditions, being a major factor determining algal local communities in Mediterranean streams (as suggested by Tornés & Sabater, 2010), that hydrological variability of each site (i.e. regardless of the flow conditions at the moment of sampling) is a factor determining local algal communities. Overall, because it has been proposed that the degree of nestedness observed in a given meta-community may be related to the processes governing species distributions across local assemblages within a region (Leibold & Mikkelson, 2002), we propose that hydrological variability is, in Mediterranean streams, a major single-dimension factor governing such processes.
Links between nestedness and species autoecology
The distinction between nested and idiosyncratic diatom species was associated with their niche attributes. As a general pattern, the idiosyncratic species were more generalist and widely distributed than the nested ones. A higher occurrence of idiosyncratic than nested species was also observed in Finish streams (Soininen, 2008; 44% versus 34% observed in this study). However, in contrast to the current study, nestedness was not related to species niche breadth in the previous study. Other studies have not observed a relationship between nestedness and niche metrics, but have measured nestedness with nested ranks rather than with T (e.g. Azeria & Kolasa, 2008). Our study included both reach- and landscape-scaled variables. This approach, combined with the fact that sites were visited twice (hence non-detected species were most probably true absences rather than ‘false zeros’), may have improved the accuracy of the OMI analyses. A positive association between T (i.e. idiosyncrasy) and common generalist species was previously proposed by Mcabendroth et al. (2005). Hence, our study confirms that the same relationship may also emerge in unicellular meta-communities.
It is interesting to note that in our study, some diatom taxa showed particularly idiosyncratic distributions and high niche breadths. Most of these species are characterised by being widespread and highly tolerant to environmental disturbances. For instance, species such as F. saprophila, N. desertorum and N. frustulum are resistant to organic pollution (Tornés et al., 2007), and others such as S. seminulum, H. veneta and N. aurariae are commonly found in highly disturbed rivers (Fore & Grafe, 2002; Rimet, 2009; Della Bella et al., 2012).
Consequences of the observed patterns for the conservation of freshwater unicellular biodiversity
In a strongly nested system, preserving a few species-rich sites would be a good strategy to preserve the regional pool of species. However, the relatively high number of idiosyncratic taxa and sites found in this study corroborates the findings that strong nestedness is rare in nature (e.g. Soininen, 2008; Sasaki et al., 2012; Ruhí et al., 2013). As intermittent streams presented the highest proportion of idiosyncratic taxa and the lowest species richness, our results suggest that in the Mediterranean region, preserving only a few species-rich sites involves a high risk of neglecting species exclusive to species-poor habitats. Moreover, intermittent sites presented the highest spatial dissimilarity, implying that each of them had particular (yet species poor) diatom communities. Thus, our findings emphasise the need to preserve a diverse network of habitats (both species rich and species poor) to maintain regional (gamma) diversity.
Furthermore, the Mediterranean region has been identified as one of the most prominent ‘hotspots’ for future climate change. It has been predicted that precipitation will decrease and surface temperature will increase during all seasons in the whole basin, with a 25% reduction in precipitation and a 4 °C warming expected during summers under the A1B and A2 scenarios by the years 2081–2100 (Giorgi & Lionello, 2008). Contrasting flow periods (high-flow and low-flow waters) are also expected to become even more pronounced (Giorgi & Lionello, 2008). Increases in the frequency and severity of floods and droughts have already been described to affect the physical, chemical and biological properties of freshwater systems negatively and accompany other changes related to human pressure (Sabater & Tockner, 2010; Timoner et al., 2012; Tornés et al., 2013). Our results imply that idiosyncratic diatom species are likely to be favoured by future global change in Mediterranean rivers and streams. Thus, the predicted scenario of lower precipitation in rivers in the Mediterranean region may lead to a loss of biodiversity and ecosystem functions.
Finally, our findings suggest that conservation strategies may become logistically more difficult, and financially very expensive, in the long term. As the current need to protect a diverse network of sites will be even more necessary in the near future (because local communities are probably to become taxonomically and functionally poorer as well as less spatially ordered), it will be more difficult to implement efficient programmes for biodiversity conservation at the regional scale. On the whole, a better understanding of how hydrological stability affects freshwater diatom communities will anticipate ecological problems under scenarios of global change and thus improve the existing tools for their conservation.
Acknowledgments
We thank Sergi Sabater for his constructive comments on the manuscript. This study was funded by the project SCARCE (Consolider-Ingenio 2010, CSD2009-00065).

