


Linking carbon and nitrogen metabolism to depth distribution of submersed macrophytes using high ammonium dosing tests and a lake survey
Summary
- Strategies of carbon (C) and nitrogen (N) utilisation are among the factors determining plant distribution. It has been argued that submersed macrophytes adapted to lower light environments are more efficient in maintaining C metabolic homeostasis due to their conservative C strategy and ability to balance C shortage. We studied how depth distributions of 12 submersed macrophytes in Lake Erhai, China, were linked to their C-N metabolic strategies when facing acute
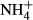 dosing.
dosing. 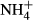 dosing changed C-N metabolism significantly by decreasing the soluble carbohydrate (SC) content and increasing the
dosing changed C-N metabolism significantly by decreasing the soluble carbohydrate (SC) content and increasing the 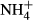 -N and free amino acid (FAA) content of plant tissues.
-N and free amino acid (FAA) content of plant tissues.- The proportional changes in SC contents in the leaves and FAA contents in the stems induced by
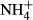 dosing were closely correlated (positive for SC and negative for FAA) with the colonising water depths of the plants in Lake Erhai, the plants adapted to lower light regimes being more efficient in maintaining SC and FAA homeostasis.
dosing were closely correlated (positive for SC and negative for FAA) with the colonising water depths of the plants in Lake Erhai, the plants adapted to lower light regimes being more efficient in maintaining SC and FAA homeostasis. - These results indicate that conservative carbohydrate metabolism of submersed macrophytes allowed the plants to colonise greater water depths in eutrophic lakes, where low light availability in the water column diminishes carbohydrate production by the plants.
Introduction
The metabolism of carbon (C) and nitrogen (N) by plants is strongly affected by environmental conditions (Myers & Kitajima, 2007; Valladares & Niinemets, 2008). The carbon gain hypothesis proposes a trade-off between plant growth rate and survival, depending on light regime (Kobe et al., 1995; Valladares & Niinemets, 2008). At low light, shade-tolerant plants increase their survival by minimising C loss, for example by having low respiration and high carbohydrate storage (Kobe, 1997; Walters & Reich, 1999; Myers & Kitajima, 2007), while shade-intolerant plants typically exhibit high growth rates and high respiration rates (Niinemets, 1997; Walters & Reich, 1999). Several studies have shown that the conservative rather than the consumptive C strategy benefits plant survival in low light environments (Thompson, Kriedemann & Craig, 1992; Gaucher et al., 2005).
Eutrophication changes light and nutrient availability in the water column, thereby affecting the distribution of submersed macrophytes in lakes (Barko & Smart, 1981; Chambers & Kalff, 1985; Middelboe & Markager, 1997). Rosette macrophytes have a lower light compensation point for photosynthesis (PI) than canopy-forming macrophytes (Titus & Adams, 1979; Su et al., 2004) and tend to be efficient at capturing CO2 from the sediment (Bowes, 1987). Canopy-forming macrophytes are capable of elongating their shoots and thereby augment their potential to reach the water surface and access atmospheric CO2 (Titus & Adams, 1979; Ni, 2001; Lucassen et al., 2009), enabling them to become dominant in eutrophic lakes. It has been argued that submersed macrophytes adapted to lower light environments are more efficient at maintaining C metabolic homeostasis due to their conservative C strategy and ability to balance C shortage (Cao et al., 2011).
Excessive ammonium (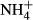 ) enrichment in the water column, for instance due to eutrophication, affects the C-N metabolism of submersed macrophytes by decreasing soluble carbohydrate (SC) content and increasing the content of NH4-N and free amino acids (FAA) in the tissues, which may be detrimental to the plants (Smolders et al., 1996; Cao, Ni & Xie, 2004; Cao et al., 2009a,b, 2011). Submersed macrophytes cannot prevent absorption of
) enrichment in the water column, for instance due to eutrophication, affects the C-N metabolism of submersed macrophytes by decreasing soluble carbohydrate (SC) content and increasing the content of NH4-N and free amino acids (FAA) in the tissues, which may be detrimental to the plants (Smolders et al., 1996; Cao, Ni & Xie, 2004; Cao et al., 2009a,b, 2011). Submersed macrophytes cannot prevent absorption of 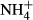 from the surrounding water (Van Katwijk et al., 1997; Britto et al., 2001). However, to avoid
from the surrounding water (Van Katwijk et al., 1997; Britto et al., 2001). However, to avoid 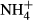 accumulation in their tissue, the plants incorporate
accumulation in their tissue, the plants incorporate 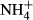 into nitrogenous organic compounds instead or transporting it out of the tissue, processes that both are energy and thus carbohydrate demanding (Britto et al., 2001; Britto & Kronzucker, 2002; Cao et al., 2009b). Experiments have demonstrated that many submersed macrophytes are able to adapt their metabolism to moderate
into nitrogenous organic compounds instead or transporting it out of the tissue, processes that both are energy and thus carbohydrate demanding (Britto et al., 2001; Britto & Kronzucker, 2002; Cao et al., 2009b). Experiments have demonstrated that many submersed macrophytes are able to adapt their metabolism to moderate 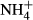 dosing by counteracting the loss of carbohydrates for
dosing by counteracting the loss of carbohydrates for 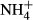 assimilation and thereby maintain a relatively stable SC content despite significant changes in FAA and starch contents and plant growth (Cao et al., 2007, 2011; Li, Cao & Ni, 2007), the latter indicating that
assimilation and thereby maintain a relatively stable SC content despite significant changes in FAA and starch contents and plant growth (Cao et al., 2007, 2011; Li, Cao & Ni, 2007), the latter indicating that 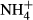 serves as a N nutrition for the plants. Probably as a result of the dual roles of
serves as a N nutrition for the plants. Probably as a result of the dual roles of 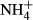 acting as N nutrition and/or toxin, the biomass of the submersed macrophyte Vallisneria natans increased at moderate
acting as N nutrition and/or toxin, the biomass of the submersed macrophyte Vallisneria natans increased at moderate  enrichment (NH4-N < 0.3 mg L−1) in the water column, but decreased at high concentrations (NH4-N > 0.56 mg L−1) in lakes along the mid-low reaches of the Yangtze River (Cao et al., 2007).
enrichment (NH4-N < 0.3 mg L−1) in the water column, but decreased at high concentrations (NH4-N > 0.56 mg L−1) in lakes along the mid-low reaches of the Yangtze River (Cao et al., 2007).
Different submersed macrophytes show different C-N metabolic responses to light availability and/or 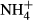 dosing (Cao et al., 2011), and the C-N metabolic responses to
dosing (Cao et al., 2011), and the C-N metabolic responses to 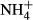 dosing are strongly dependent on light availability (Zhang et al., 2010; Cao et al., 2011). It is reasonable to assume then that there is a relationship between the depth distributions of the various submersed macrophytes and their C-N metabolic strategies when faced with
dosing are strongly dependent on light availability (Zhang et al., 2010; Cao et al., 2011). It is reasonable to assume then that there is a relationship between the depth distributions of the various submersed macrophytes and their C-N metabolic strategies when faced with 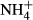 dosing. To explore this further, we conducted a field survey of the depth distributions and biomass of submerged macrophytes as well as
dosing. To explore this further, we conducted a field survey of the depth distributions and biomass of submerged macrophytes as well as 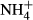 dosing experiments to test how high
dosing experiments to test how high 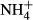 dosing affects the C-N metabolism of different forms of submersed macrophytes and specifically to test the hypothesis that submersed macrophytes adapted to a lower light environment have a conservative C-N metabolic strategy in response to
dosing affects the C-N metabolism of different forms of submersed macrophytes and specifically to test the hypothesis that submersed macrophytes adapted to a lower light environment have a conservative C-N metabolic strategy in response to  dosing.
dosing.
Methods
Field survey of submersed macrophytes
A field survey of the depth distribution of submersed macrophytes was carried out in September 2009 in large mesotrophic Lake Erhai (25°52′N, 100°06′E) located in the Yunnan Province, China. The lake has a surface area of 250 km2 and a mean water depth of 11 m (maximum depth 21 m). Total phosphorus (TP) and total nitrogen (TN) concentrations in the water column amounted to, respectively, 0.03 mg L−1 and 0.7 mg L−1, and Secchi transparency was 1.5 m. Submersed macrophytes covered 40% of the lake surface in the 1980s, but less than 8% in 2009 due to eutrophication (Dai, 1984; Li et al., 2011).
Submersed macrophytes were sampled in triplicate at intervals of 0.5-m water depths at sites along transects from the shore to the deepest depths where the plants occurred. A total of 844 samples were collected from 79 transects uniformly distributed around the lakeshore. At transects with a steep lake bottom, submersed macrophytes were sampled at fewer sites. The macrophytes were collected by an underwater reaping hook, covering a bottom surface area of 0.25 m2, and subsequently weighed to determine the fresh biomass (FW) of each species. At each site, underwater photosynthetic available radiation (PAR) was measured just beneath the water surface and at depths of 0.5 and 1.5 m by a radiation sensor (UW-192) connected to a data logger (Li-1400; Li-cor Company, Lincoln, NE, U.S.A), and water depth was recorded. The attenuation coefficient (K) in the water column was calculated based on PAR at different depths. PAR at the bottom was calculated as IZ = I0 (1-KZ)/(KZ), where IZ is irradiance in depth of Z m, I0 is irradiance in water surface (Duarte, Kalff & Peters, 1986) and expressed as a percentage of light intensity relative to the level just beneath the water surface. Ten submersed macrophytes, Potamogeton pectinatus, Potamogeton maackianus, Potamogeton lucens, Potamogeton perfoliatus, Potamogeton intortifolius, Potamogeton malaianus, Myriophyllum spicatum, Vallisneria natans, Ceratophyllum demersum and Hydrilla verticillata, were found in this survey (Diao, 1990; Sun, 1992).
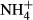 dosing experiment
dosing experiment
An experiment was conducted outdoors in Xizhou town near to the shore of Lake Erhai from 26 April to 13 May 2011. Twelve submersed macrophytes species (including the 10 macrophyte species found in the survey, and the less common Potamogeton acutifolius and Potamogeton crispus in Lake Erhai but not recorded in the present survey) were used to examine C-N metabolic responses to 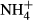 dosing. Seedlings (15 cm height) of V. natans and apical shoots (15 cm length) of the other plants were collected from the lake at depths ranging from 0.5 to 2 m, transplanted into 576 cylinder pots (diameter: 6.0 cm, height: 7.5 cm; 48 pots per species) containing 5-cm lake sediment and then placed uniformly in 96 tanks (27 × 20 × 30 cm height; 8 tanks per species) under a roof shading out about 50% of the sunlight. The plants were kept for a 15-day acclimation by filling the tanks with the lake water. During this period, the water in each tank was renewed and filamentous algae on plant leaves were gently removed every 5 days. PAR at 5 cm below the water surface was approximately 500 μmol m−2 s−1 at noon. Water temperature ranged from 16.3 to 23.8 °C. The concentrations of NO3-N, NH4-N and PO4-P in the water column were 0.39, 0.02 and 0.005 mg L−1, respectively, and did not change greatly during the acclimation period. The sediment was collected about 1 Km offshore in Lake Erhai, and the concentrations of TN, TP and organic material in the sediments were 2.12 mg g−1, 0.68 mg g−1 and 25.6 mg g−1, respectively.
dosing. Seedlings (15 cm height) of V. natans and apical shoots (15 cm length) of the other plants were collected from the lake at depths ranging from 0.5 to 2 m, transplanted into 576 cylinder pots (diameter: 6.0 cm, height: 7.5 cm; 48 pots per species) containing 5-cm lake sediment and then placed uniformly in 96 tanks (27 × 20 × 30 cm height; 8 tanks per species) under a roof shading out about 50% of the sunlight. The plants were kept for a 15-day acclimation by filling the tanks with the lake water. During this period, the water in each tank was renewed and filamentous algae on plant leaves were gently removed every 5 days. PAR at 5 cm below the water surface was approximately 500 μmol m−2 s−1 at noon. Water temperature ranged from 16.3 to 23.8 °C. The concentrations of NO3-N, NH4-N and PO4-P in the water column were 0.39, 0.02 and 0.005 mg L−1, respectively, and did not change greatly during the acclimation period. The sediment was collected about 1 Km offshore in Lake Erhai, and the concentrations of TN, TP and organic material in the sediments were 2.12 mg g−1, 0.68 mg g−1 and 25.6 mg g−1, respectively.
In the 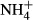 treatments, ammonium chloride solution was added to the 48 tanks (4 tanks per species) at noon to obtain a concentration of 3 mg L−1 NH4-N, and 48 h later, the plants were collected for analysis of C-N metabolites. The tanks without
treatments, ammonium chloride solution was added to the 48 tanks (4 tanks per species) at noon to obtain a concentration of 3 mg L−1 NH4-N, and 48 h later, the plants were collected for analysis of C-N metabolites. The tanks without 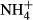 supplement were regarded as the controls. Plants collected from the same tank were washed with distilled water three times and then carefully separated into leaves, stems and roots, dried with tissue paper and stored at −20 °C. The frozen plants were ground into fine powder in liquid nitrogen with a mortar and a pestle. About 0.5-g sample powder was extracted twice with 8 ml 80% ethanol at 80 °C for 20 min and then centrifuged at 10 000 × g for 15 min. The supernatant was collected, decolourised by activated charcoal and filtered (microvoid filter film, φ20 mm). The filtrate was used for analysis of NH4-N, SC and FAA (Yemm & Willis, 1954; Yemm, Cocking & Ricketts, 1955; Hecht & Mohr, 1990) using ammonium sulphate, glucose and alanine as standards, respectively. The residue after centrifugation was used for analysis of starch (Dirk et al., 1999). Contents of C-N metabolites in the leaves and the stems were expressed as mg g−1 dry weight by multiplying their fresh to dry weight ratios.
supplement were regarded as the controls. Plants collected from the same tank were washed with distilled water three times and then carefully separated into leaves, stems and roots, dried with tissue paper and stored at −20 °C. The frozen plants were ground into fine powder in liquid nitrogen with a mortar and a pestle. About 0.5-g sample powder was extracted twice with 8 ml 80% ethanol at 80 °C for 20 min and then centrifuged at 10 000 × g for 15 min. The supernatant was collected, decolourised by activated charcoal and filtered (microvoid filter film, φ20 mm). The filtrate was used for analysis of NH4-N, SC and FAA (Yemm & Willis, 1954; Yemm, Cocking & Ricketts, 1955; Hecht & Mohr, 1990) using ammonium sulphate, glucose and alanine as standards, respectively. The residue after centrifugation was used for analysis of starch (Dirk et al., 1999). Contents of C-N metabolites in the leaves and the stems were expressed as mg g−1 dry weight by multiplying their fresh to dry weight ratios.
Statistical analysis
SPSS software was used for statistical analyses. All data were tested for normality using the Shapiro–Wilk test and homogeneity using Levene's test. For each species, biomass, colonising depth and PAR requirement at the bottom were given as average values for the sites where the plant was collected. The proportional changes in NH4-N, FAA, SC and starch concentrations in the plants exposed to 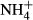 dosing (subscript F) were compared with those of the controls (subscript C), indicated as NH4-N(F-C)/C, FAA(F-C)/C, SC(F-C)/C and starch(F-C)/C, respectively. An independent t-test was used to evaluate the effects of
dosing (subscript F) were compared with those of the controls (subscript C), indicated as NH4-N(F-C)/C, FAA(F-C)/C, SC(F-C)/C and starch(F-C)/C, respectively. An independent t-test was used to evaluate the effects of 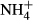 dosing on C-N metabolite contents. Spearman's rank correlation analysis was performed to compare the C-N metabolite contents and C-N metabolic indices (NH4-N, FAA, SC, starch, NH4-N(F-C)/C, FAA(F-C)/C, SC(F-C)/C and starch(F-C)/C) as well as biomass, colonising depth and PAR requirement among the 12 species.
dosing on C-N metabolite contents. Spearman's rank correlation analysis was performed to compare the C-N metabolite contents and C-N metabolic indices (NH4-N, FAA, SC, starch, NH4-N(F-C)/C, FAA(F-C)/C, SC(F-C)/C and starch(F-C)/C) as well as biomass, colonising depth and PAR requirement among the 12 species.
Results
Carbon and nitrogen metabolism of the submersed macrophytes in response to acute 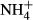 dosing
dosing
The contents of C-N metabolites differed markedly among the 12 macrophytes. In the control, the average contents of NH4-N, FAA, SC and starch of the 12 macrophytes were, respectively, 0.23 (0.07–0.84) mg g−1, 4.37 (1.57–8.52) mg g−1, 94 (40–166) mg g−1 and 66 (6.6–253) mg g−1 in the leaves and 0.34 (0.07–1.09) mg g−1, 10.6 (2.45–33.2) mg g−1, 107 (30–206) mg g−1 and 249 (35.3–502) mg g−1 in the stems (Fig. 1).

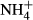 dosing experiments. The values were mean + SE (n = 4) and dry weight basis. Significant effect of
dosing experiments. The values were mean + SE (n = 4) and dry weight basis. Significant effect of 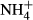 dosing is indicated by asterisk (*P < 0.05, **P < 0.01, ***P < 0.001). The species on the x-axis were ordered in a decreasing rank according to SC(F-C)/C in the leaves. Note different scales on the y axis.
dosing is indicated by asterisk (*P < 0.05, **P < 0.01, ***P < 0.001). The species on the x-axis were ordered in a decreasing rank according to SC(F-C)/C in the leaves. Note different scales on the y axis.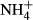 dosing increased NH4-N and FAA concentrations and decreased SC concentrations in the leaves and the stems of all macrophyte species except for the SC concentrations in the stems of P. pectinatus, and decreased or increased the starch contents in the plant tissue, depending on plant species. The NH4-N(F-C)/C, FAA(F-C)/C, SC(F-C)/C and starch(F-C)/C were, respectively, 2.89 to 27.7, 1.52 to 17.1, −0.81 to −0.22 and −0.76 to 1.06 in the leaves and 0.10 to 4.21, 0.34 to 2.51, −0.66 to 0.11 and −0.49 to 0.40 in the stems. In general, the responses of C-N metabolites to
dosing increased NH4-N and FAA concentrations and decreased SC concentrations in the leaves and the stems of all macrophyte species except for the SC concentrations in the stems of P. pectinatus, and decreased or increased the starch contents in the plant tissue, depending on plant species. The NH4-N(F-C)/C, FAA(F-C)/C, SC(F-C)/C and starch(F-C)/C were, respectively, 2.89 to 27.7, 1.52 to 17.1, −0.81 to −0.22 and −0.76 to 1.06 in the leaves and 0.10 to 4.21, 0.34 to 2.51, −0.66 to 0.11 and −0.49 to 0.40 in the stems. In general, the responses of C-N metabolites to 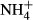 dosing were NH4-N > FAA > SC > starch in both the leaves and the stems, and the C-N metabolites in the leaves were more responsive to
dosing were NH4-N > FAA > SC > starch in both the leaves and the stems, and the C-N metabolites in the leaves were more responsive to 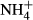 dosing than those in the stems.
dosing than those in the stems.
In the leaves, the NH4-N content was positively correlated with FAA and negatively with SC, and FAA negatively with SC (P < 0.001 for all). In the stems, NH4-N, FAA and starch contents were positively correlated with each other (P < 0.01 for all), and SC contents were positively correlated with starch (P = 0.002). Also, NH4-N and FAA in the leaves correlated positively with NH4-N, FAA and SC in the stems (Table 1; P < 0.01 for all).
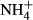 dosing experiments. (n = 96)
dosing experiments. (n = 96)| NH4-N | FAA | SC | Starch | |
|---|---|---|---|---|
| In the leaves | ||||
| FAA | 0.92 (<0.001) | |||
| SC | −0.43 (<0.001) | −0.36 (<0.001) | ||
| Starch | 0.06 (0.579) | 0.07 (0.499) | 0.16 (0.111) | |
| In the stems | ||||
| FAA | 0.92 (<0.001) | |||
| SC | 0.09 (0.396) | 0.05 (0.633) | ||
| Starch | 0.27 (0.007) | 0.45 (<0.001) | 0.31 (0.002) | |
| Between the leaves and the stems | ||||
| NH4-N in leaf | 0.57 (<0.001) | 0.66 (<0.001) | 0.06 (0.583) | −0.09 (0.360) |
| FAA in leaf | 0.54 (<0.001) | 0.66 (<0.001) | 0.06 (0.586) | −0.03 (0.754) |
| SC in leaf | −0.32 (0.002) | −0.28 (0.006) | 0.73 (<0.001) | −0.26 (0.012) |
| Starch in leaf | −0.04 (0.670) | 0.08 (0.418) | 0.40 (<0.001) | 0.28 (0.005) |
- Bold indicates P < 0.05.
When plotting leaf SC(F-C)/C against stem FAA(F-C)/C, the macrophytes with C-N metabolism most responsive to 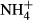 dosing (P. perfoliatus, P. intortifolius, P. malaianus, P. acutifolius and P. crispus) were grouped in the upper left side of the coordinate plane, while the macrophytes with C-N metabolism less sensitive to
dosing (P. perfoliatus, P. intortifolius, P. malaianus, P. acutifolius and P. crispus) were grouped in the upper left side of the coordinate plane, while the macrophytes with C-N metabolism less sensitive to 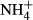 dosing (M. spicatum, P. pectinatus, C. demersum, P. maackianus and V. natans) were grouped in the lower right side (Fig. 2).
dosing (M. spicatum, P. pectinatus, C. demersum, P. maackianus and V. natans) were grouped in the lower right side (Fig. 2).

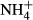 dosing. Cd: C. demersum, Ms: M. spicatum, Ppc: P. pectinatus, Vn: V. natans, Pl: P. lucens, Pml: P. malaianus, Pi: P. intortifolius, Ppf: P. perfoliatus, Pmk: P. maackianus, Pa: P. acutifolius, Pc: P. crispus and Hv: H. verticillata.
dosing. Cd: C. demersum, Ms: M. spicatum, Ppc: P. pectinatus, Vn: V. natans, Pl: P. lucens, Pml: P. malaianus, Pi: P. intortifolius, Ppf: P. perfoliatus, Pmk: P. maackianus, Pa: P. acutifolius, Pc: P. crispus and Hv: H. verticillata. Relationships between C-N metabolism of submersed macrophytes in acute 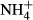 dosing and their colonising water depth in the lake
dosing and their colonising water depth in the lake
In the field investigation, biomass, colonising depths and PAR requirements of the 10 macrophyte species were 0.77–4.56 kg FW m−2, 1.70–3.55 m and 2.7–14.7%, respectively, with P. maackianus colonising the deepest water and exhibiting the highest biomass, while P. intortifolius colonised the shallowest water and had the lowest biomass. Across the 10 macrophyte species, SC(F-C)/C in the leaves correlated positively with colonising depth (r = 0.89, P = 0.001) and biomass (r = 0.85, P = 0.002) and negatively with PAR requirements (r = −0.82, P = 0.004); FAA(F-C)/C in the stems correlated negatively with colonising depth (r = −0.82, P = 0.004), positively with PAR requirements (r = 0.73, P = 0.016) and was marginally negatively related to biomass (r = −0.55, P = 0.098; Fig. 3). The colonising depth was negatively correlated with NH4-N(F-C)/C in the stems (r = −0.65, P = 0.043). However, no significant relationship was found between colonising depth, PAR requirement and biomass and the NH4-N, FAA, SC and starch contents and NH4-N(F-C)/C and starch(F-C)/C in the leaves and the stems (Table 2).
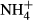 dosing (n = 10)
dosing (n = 10)| Colonising depth (m) | PAR requirements (%) | Biomass (g m−2) | |
|---|---|---|---|
| Proportions in the leaves | |||
| NH4-N(F-C)/C | −0.27 (0.446) | 0.15 (0.676) | −0.52 (0.128) |
| FAA(F-C)/C | −0.52 (0.128) | 0.35 (0.328) | −0.68 (0.029) |
| SC(F-C)/C | 0.89 (0.001) | −0.82 (0.004) | 0.85 (0.002) |
| Starch(F-C)/C | 0.21 (0.556) | −0.02 (0.960) | 0.21 (0.556) |
| Proportions in the stems | |||
| NH4-N(F-C)/C | −0.65 (0.043) | 0.49 (0.150) | −0.33 (0.347) |
| FAA(F-C)/C | −0.82 (0.004) | 0.73 (0.016) | −0.55 (0.098) |
| SC(F-C)/C | 0.25 (0.489) | −0.21 (0.556) | 0.10 (0.777) |
| Starch(F-C)/C | 0.16 (0.651) | −0.01 (0.987) | 0.21 (0.556) |
- Bold indicates P < 0.05.

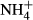 dosing, respectively.
dosing, respectively.Discussion
We found that the submersed macrophytes adapted to lower light environments were more efficient at maintaining C-N metabolic homeostasis when faced with acute 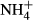 dosing. The close relationships between SC(F-C)/C in leaves and colonising water depths, PAR at the bottom and biomass of the plants in Lake Erhai indicate that growth, survival and distribution of the plants in this lake and probably also in other eutrophic lakes are closely linked to carbohydrate metabolism. In plant leaves, 2-oxoglutarate acts as a precursor for
dosing. The close relationships between SC(F-C)/C in leaves and colonising water depths, PAR at the bottom and biomass of the plants in Lake Erhai indicate that growth, survival and distribution of the plants in this lake and probably also in other eutrophic lakes are closely linked to carbohydrate metabolism. In plant leaves, 2-oxoglutarate acts as a precursor for 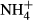 assimilation and is derived largely from carbon skeletons generated by dark respiration (Lasa et al., 2002; Gauthier et al., 2010). The proportional changes in SC contents induced by the acute
assimilation and is derived largely from carbon skeletons generated by dark respiration (Lasa et al., 2002; Gauthier et al., 2010). The proportional changes in SC contents induced by the acute 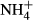 dosing in our study therefore possibly mirror the relative proportion of carbon skeleton production generated by dark respiration to total carbohydrate production in the plants.
dosing in our study therefore possibly mirror the relative proportion of carbon skeleton production generated by dark respiration to total carbohydrate production in the plants.
We found low carbohydrate contents in C. demersum, M. spicatum and V. natans, which typically inhabit eutrophic lakes (Chambers, 1987; Chambers & Kalff, 1987; Cao et al., 2011) where light availability is low. These species are therefore expected to have a carbohydrate metabolism similar to shade-tolerant plants, characterised by low rates of photosynthesis and respiration (Givnish, 1988; Noguchi, Sonoike & Terashima, 1996; Noguchi et al., 2001). Further evidence was derived from a 2-month experiment with these three species run at contrasting light (27 versus 125 μmol m−2s−1) and 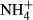 (0.03 versus 0.21 mg L−1 NH4-N) environments (Cao et al., 2011), showing relatively stable SC contents of the plants even though the growth and FAA contents changed greatly. Similarly, SC contents have been reported to be relatively unresponsive to varying light regimes in terrestrial shade-tolerant plants as compared with sun plants (Noguchi et al., 2001; Pystina & Danilov, 2001).
(0.03 versus 0.21 mg L−1 NH4-N) environments (Cao et al., 2011), showing relatively stable SC contents of the plants even though the growth and FAA contents changed greatly. Similarly, SC contents have been reported to be relatively unresponsive to varying light regimes in terrestrial shade-tolerant plants as compared with sun plants (Noguchi et al., 2001; Pystina & Danilov, 2001).
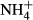 dosing increased NH4-N and FAA content and decreased SC content in leaves and stems of all the macrophytes except for the stem SC content of P. pectinatus. Similar results have been reported for other plants experiencing high
dosing increased NH4-N and FAA content and decreased SC content in leaves and stems of all the macrophytes except for the stem SC content of P. pectinatus. Similar results have been reported for other plants experiencing high 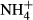 dosing (Cao et al., 2007, 2009a). In our study, the extent to which the
dosing (Cao et al., 2007, 2009a). In our study, the extent to which the 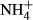 dosing changed the FAA and SC contents was species specific and also dependent on the type of organs (leaf or stem), with the plant species inhabiting shallow water (P. perfoliatus, P. intortifolius, P. malaianus, P. acutifolius and P. crispus) being more responsive to
dosing changed the FAA and SC contents was species specific and also dependent on the type of organs (leaf or stem), with the plant species inhabiting shallow water (P. perfoliatus, P. intortifolius, P. malaianus, P. acutifolius and P. crispus) being more responsive to 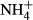 dosing than those from deep waters (M. spicatum, P. pectinatus, C. demersum, P. maackianus and V. natans). It is not clear, however, why the submersed macrophytes adapted to a high light climate (shallow waters) were more prone to accumulate FAA when faced with high
dosing than those from deep waters (M. spicatum, P. pectinatus, C. demersum, P. maackianus and V. natans). It is not clear, however, why the submersed macrophytes adapted to a high light climate (shallow waters) were more prone to accumulate FAA when faced with high 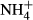 dosing, but it is known that leaves of sun plants exposed to high light need more proteins to support high photosynthesis and have high
dosing, but it is known that leaves of sun plants exposed to high light need more proteins to support high photosynthesis and have high 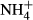 turnover in photorespiration (Givnish, 1988; Noguchi et al., 2001; Pystina & Danilov, 2001). Colonising water depths were closely correlated with SC(F-C)/C in leaves but not with FAA(F-C)/C in leaves, possibly because plants experiencing high
turnover in photorespiration (Givnish, 1988; Noguchi et al., 2001; Pystina & Danilov, 2001). Colonising water depths were closely correlated with SC(F-C)/C in leaves but not with FAA(F-C)/C in leaves, possibly because plants experiencing high 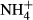 dosing lost much more carbohydrate than the amount of carbon skeletons for incorporating
dosing lost much more carbohydrate than the amount of carbon skeletons for incorporating 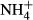 into FAA (Britto et al., 2001; Cao et al., 2004, 2009a), which might have weakened a relationship between colonising water depths and FAA(F-C)/C in comparison with the relationship for the SC(F-C)/C in leaves. We can exclude the possibility that the differences in colonising water depths among the macrophytes were due to depth-dependent
into FAA (Britto et al., 2001; Cao et al., 2004, 2009a), which might have weakened a relationship between colonising water depths and FAA(F-C)/C in comparison with the relationship for the SC(F-C)/C in leaves. We can exclude the possibility that the differences in colonising water depths among the macrophytes were due to depth-dependent 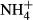 toxicity in Lake Erhai, because its lake water is fully mixed at all depths and
toxicity in Lake Erhai, because its lake water is fully mixed at all depths and 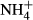 concentrations were low in the lake water (Yang et al., 2012) and below the toxic
concentrations were low in the lake water (Yang et al., 2012) and below the toxic 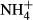 levels reported for submersed macrophytes (Van Katwijk et al., 1997; Smolders, Lucassen & Roelofs, 2002; Cao et al., 2007) and far below the
levels reported for submersed macrophytes (Van Katwijk et al., 1997; Smolders, Lucassen & Roelofs, 2002; Cao et al., 2007) and far below the 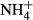 concentration in our experiment.
concentration in our experiment.
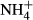 dosing induced FAA accumulation in both the leaves and the stems of the macrophytes, but FAA contents were lower in the leaves than the stems, indicating that transportation of FAA from leaves to stems was not mediated by passive but rather by active transport, which consumes energy and relates to plant basic metabolic strength, with shade-tolerant plants spending less energy in transporting metabolites than sun plants ((Noguchi et al., 2001). Given that stems of submersed macrophytes act as an N sink in response to
dosing induced FAA accumulation in both the leaves and the stems of the macrophytes, but FAA contents were lower in the leaves than the stems, indicating that transportation of FAA from leaves to stems was not mediated by passive but rather by active transport, which consumes energy and relates to plant basic metabolic strength, with shade-tolerant plants spending less energy in transporting metabolites than sun plants ((Noguchi et al., 2001). Given that stems of submersed macrophytes act as an N sink in response to 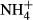 fertilisation (Invers et al., 2004), the differences in harvesting light and producing energy among the plant species tested in our study might have affected FAA transportation and FAA(F-C)/C in stems of the plants, leading to the observed close correlation of FAA(F-C)/C in stems and SC(F-C)/C in leaves with colonising depths and light requirements of the different species. No significant relationship was observed between colonising depth, PAR requirement and biomass and the NH4-N, FAA, SC and starch contents and NH4-N(F-C)/C and starch(F-C)/C in the leaves and the stems, possibly because these metabolic indices are not so directly linked to
fertilisation (Invers et al., 2004), the differences in harvesting light and producing energy among the plant species tested in our study might have affected FAA transportation and FAA(F-C)/C in stems of the plants, leading to the observed close correlation of FAA(F-C)/C in stems and SC(F-C)/C in leaves with colonising depths and light requirements of the different species. No significant relationship was observed between colonising depth, PAR requirement and biomass and the NH4-N, FAA, SC and starch contents and NH4-N(F-C)/C and starch(F-C)/C in the leaves and the stems, possibly because these metabolic indices are not so directly linked to 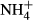 assimilation and FAA transportation as are SC and FAA (Mark, 1999; Britto & Kronzucker, 2005).
assimilation and FAA transportation as are SC and FAA (Mark, 1999; Britto & Kronzucker, 2005).
To conclude, we found relationships between the distribution of submersed macrophytes and their C-N metabolic responses to acute 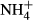 dosing and also that a conservative strategy of carbohydrate metabolism in response to
dosing and also that a conservative strategy of carbohydrate metabolism in response to  dosing allowed some species to colonise deeper waters. To our knowledge, this is the first time that an
dosing allowed some species to colonise deeper waters. To our knowledge, this is the first time that an 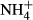 dosing test has been used to explore C-N metabolism of submersed macrophytes adapted to various light climates. It should be noted that
dosing test has been used to explore C-N metabolism of submersed macrophytes adapted to various light climates. It should be noted that 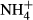 may act as N nutrition and/or be toxic to submersed macrophytes (Britto & Kronzucker, 2002), and it has been shown that the effects of
may act as N nutrition and/or be toxic to submersed macrophytes (Britto & Kronzucker, 2002), and it has been shown that the effects of 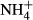 dosing on C-N metabolism of submersed macrophytes are dependent on
dosing on C-N metabolism of submersed macrophytes are dependent on 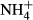 concentration, exposure time and PAR (Cao et al., 2009a,b, 2011). Therefore, long-term
concentration, exposure time and PAR (Cao et al., 2009a,b, 2011). Therefore, long-term 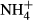 enrichment in eutrophic lakes may potentially affect C-N metabolism of submersed macrophytes in a different manner than observed in our study.
enrichment in eutrophic lakes may potentially affect C-N metabolism of submersed macrophytes in a different manner than observed in our study.
Acknowledgments
This study was supported by the National Science Foundation of China (Grant No. 31270508, 41230853) and the National High Technology Research and Development Program of China (Grant No. 2012ZX07105-004). Erik Jeppesen was supported by the EU FP-7 project REFRESH (Adaptive strategies to Mitigate the Impacts of Climate Change on European Freshwater Ecosystems, Contract No.: 244121), ‘CLEAR’ (a Villum Kann Rasmussen Centre of Excellence project) and the STF project CRES. We thank Anne Mette Poulsen for editorial comments and thank two anonymous reviewers for their valuable comments on the manuscript.

