

Evaluation of Behavior and Survival of Fish Exposed to an Axial-Flow Hydrokinetic Turbine
Abstract
Previous studies have evaluated fish injury and mortality at hydrokinetic (HK) turbines, but because these studies focused on the impacts of these turbines in situ they were unable to evaluate fish responses to controlled environmental characteristics (e.g., current velocity and light or dark conditions). In this study, we used juvenile hybrid Striped Bass (HSB; Striped Bass Morone saxatilis × White Bass M. chrysops; N = 620), Rainbow Trout Oncorhynchus mykiss (N = 3,719), and White Sturgeon Acipenser transmontanus (N = 294) in a series of laboratory experiments to (1) evaluate the ability of fish to avoid entrainment through an axial-flow HK turbine, (2) evaluate fish injury and survival associated with turbine entrainment, and (3) compare the effects of different HK turbines on fish. We found that the probability of turbine entrainment was species dependent and highest for HSB. Across species, current velocity influenced entrainment probability. Among entrained fish, observed survival rates were generally >0.95. The probability of injury for surviving entrained fish only differed from that for nonentrained fish for Rainbow Trout and in general was not >0.20. The probability of injury following entrainment was greater only for HSB, although there were no differences in injury rates between fish that were turbine entrained and those that were not, suggesting that injuries were not turbine related. Taking turbine entrainment, survival, and injury estimates together allowed us to estimate the probability of a randomly selected fish in a population proximate to an HK turbine surviving passage or remaining uninjured after passage. For species and current velocities for which there was a significant effect due to entrainment, we estimated, for instance, that HSB had a survival probability of 0.95 and that Rainbow Trout and White Sturgeon had a >0.99 probability of survival. Similarly, by combining these estimates with those from previous studies, we derived total passage survival probabilities >0.90 but generally approaching 1.00 across different HK turbine types, fish species, and fish lengths.
Received May 14, 2014; accepted October 23, 2014
There is considerable interest in the development of marine and hydrokinetic (HK) energy projects in rivers, estuaries, and coastal ocean waters around the world. Hydrokinetic technologies convert the energy of water currents into electricity without the adverse impacts of the dams and impoundments associated with conventional hydropower (which relies on the flow of water from a higher elevation to generate electricity) or the extraction and combustion of fossil fuels. In particular, the renewable energy potential in U.S. river and ocean currents is very large. For instance, the Electric Power Research Institute (EPRI 2011a, 2012) has estimated that approximately one-third of all U.S. electricity needs could be met by harnessing energy from river and ocean currents and waves. At present, however, only a small number of HK turbines have been deployed in U.S. waters, in part because the environmental impacts of these turbines are not known.
Although numerous HK designs are under development (see DOE 2009 for a description of the technologies and their potential environmental effects), the most commonly proposed projects entail arrays of rotating devices, much like submerged wind turbines, that are positioned in high-velocity (high-energy) areas such as tidal and river channels. The diversity of HK designs implies a diversity of environmental impacts (Čada et al. 2007, 2011), but a potential impact common to most designs is the risk of blade strike to aquatic organisms (Wilson et al. 2007). Only a limited number of studies have examined the risk of blade strike that HK technologies pose to freshwater fish (Turnpenny et al. 1992; NAI 2009; Amaral et al. 2011; Seitz et al. 2011; Castro-Santos and Haro 2015), and the probabilities of blade encounter and turbine rotor passage survival are unknown for most species of fish. One study (EPRI 2011b) suggested that HK turbines present less harsh physical conditions to entrained fish than conventional hydropower turbines. For example, HK turbine runners typically cause smaller changes in shear stress, turbulence, and water pressure than do conventional turbines. Specifically, the relatively open configuration of HK turbines allows fish to avoid turbine passage and, if they are unable to avoid entrainment, the slow blade rotation rate should result in less damaging strikes and better fish survival than with conventional turbines (EPRI 2011b).
An important consideration for fish and HK turbine interactions is whether fish residing in or moving through an area with turbines will ever encounter a rotor. To be killed by HK turbine passage, a fish must (1) inhabit or pass through the water body in the vicinity of the HK turbine, (2) become unavoidably entrained in the “cylinder” of water that is destined to pass through the HK turbine, (3) be struck by a rotor blade (strike probability will primarily depend on fish behavior and length, the number of blades and their rotational speed, and the angle of the flow entering the blade-swept area), and (4) receive lethal injuries if struck by a blade (a function of leading-edge blade thickness, fish length, and relative velocity of the fish to that of the blade). With regard to the first condition, the probability of encountering a midwater turbine will be low for particular species or life stages that typically move downstream near shore, on the bottom, or near the surface of a river or tidal area (Čada and Bevelhimer 2011; Schweizer et al. 2011). Incorporation of habitat preference–based encounter probabilities into the analysis may lead to higher total passage survival than would be obtained using only turbine survival and avoidance probabilities. Encounter probabilities can be derived from information on species habitat preferences and migratory patterns. Tracking tagged fish approaching and passing turbines in field situations using radio- or acoustic telemetry techniques or bottom-mounted sonar can provide this information if it does not already exist.
Most of the elements that influence HK turbine survival (i.e., HK encounter and entrainment probabilities, HK rotor avoidance by entrained fish, and rotor blade strike mortality) are difficult to quantify in a field setting. To better understand the probabilities of HK turbine passage survival, we conducted a series of tests in a large laboratory flume that quantified (1) the ability of fish to avoid HK rotor passage and (2) blade strike injuries and mortalities that occurred when fish were forced to pass through the HK rotor. We then used this information to (3) calculate the probability of entrainment and survival given avoidance behaviors and (4) compare estimates of passage survival among HK turbine types from our own and previous studies. The tests were carried out using a ducted axial-flow HK turbine and three fish species with differing morphologies and behaviors. The ability of these fish to avoid HK turbine passage was evaluated under both light and dark conditions.
METHODS
Biological testing was conducted with a ducted, axial-flow HK turbine developed by Free Flow Power (FFP; Boston, Massachusetts). The turbine is 1.5 m in diameter and has seven blades (Figure 1). The expected rotational speeds for this unit range from 40 to 125 rpm at stream flow velocities of 1–3 m/s. This rotational speed is slower than those of many similarly sized conventional (i.e., Francis and Kaplan) turbines (EPRI 2011c). Biological evaluations included two types of tests, one designed to assess behavioral interactions and avoidance as fish moved downstream and encountered the turbine and another designed to estimate turbine passage survival. Our experimental protocols and pre- and posttest holding conditions and observations are summarized here and provided in detail in Amaral et al. (2014).

Schematic in-plan view of the test flume showing the locations of the FFP turbine, the enclosure net, and the fish injection pipes used for the turbine passage survival trials. Water flow is from left to right.
Study Species
Interactions of fish with the FFP turbine were evaluated for three species: hybrid Striped Bass (HSB; Striped Bass Morone saxatilis × White Bass M. chrysops), Rainbow Trout Oncorhynchus mykiss, and White Sturgeon Acipenser transmontanus (Table 1). These species represent different families of fish that will be susceptible to HK turbine entrainment in large rivers but that have different morphologies and that were expected to exhibit different avoidance behaviors and sensitivities to blade strike injuries. Rainbow Trout are a widely distributed sport fish that are considered representative of other trout and salmon species in terms of swimming behavior and, with respect to blade strike injury and survival, are similar to many other teleost species (i.e., bony fishes common to riverine and estuarine environments). White Sturgeon are considered representative of other sturgeon species that are likely to encounter HK turbines in rivers and the nearshore ocean. Also, evidence from blade-strike and turbine passage studies indicates that sturgeons are less susceptible to injury from blade strikes than are teleost species. There may also be behavioral differences that lead sturgeons to react differently to HK turbines than trouts and other common teleost fishes. Hybrid Striped Bass have a different body shape and are typically considered to be weaker swimmers at higher velocities than Rainbow Trout and White Sturgeon (Mellas and Haynes 1985).
| Species | Water velocity (m/s) | Mean FL (mm) (SD) | No. of trials | Test group | Live recoveries (1 h) | Dead recoveriesa (1 h) | Dead recoveriesa (48 h) |
|---|---|---|---|---|---|---|---|
| Hybrid Striped Bass | 1.5 | 131 (30.1) | 3 | T | 145 | 3 | 12 |
| C | 148 | 0 | 2 | ||||
| 2.0 | 118 (25.7) | 3 | T | 146 | 0 | 6 | |
| C | 151 | 1 | 6 | ||||
| Rainbow Trout | 1.5 (small) | 172 (12.8) | 5 | T | 404 | 0 | 0 |
| C | 500 | 0 | 0 | ||||
| 2.0 (small) | 168 (15.5) | 5 | T | 477 | 3 | 3 | |
| C | 477 | 1 | 0 | ||||
| 1.5 (large) | 271 (21.6) | 5 | T | 392 | 0 | 0 | |
| C | 471 | 0 | 0 | ||||
| 2.0 (large) | 246 (16.5) | 5 | T | 478 | 11 | 1 | |
| C | 501 | 0 | 0 | ||||
| White Sturgeon | 1.5 | 123 (14.7) | 1 | T | 69 | 0 | 1 |
| C | 74 | 0 | 1 | ||||
| 2.0 | 126 (14.6) | 1 | T | 74 | 0 | 0 | |
| C | 76 | 0 | 0 |
- a All treatment fish recovered dead downstream of the turbine at the end of a trial were assumed to be turbine-passed fish.
Striped Bass are an anadromous species that could be expected to encounter HK turbines in both rivers and tidal streams. We expected HSB and White Sturgeon to have the highest probabilities of entrainment due to their lesser burst swimming capabilities compared with Rainbow Trout (Mellas and Haynes 1985). We also expected that due to their tough body armoring White Sturgeon would have a lower probability of injury and mortality than HSB and Rainbow Trout.
Test Turbine and Flume Design
The FFP turbine was installed in a large, recirculating laboratory flume at the Alden Research Laboratory in Holden, Massachusetts (Figure 1). The test flume has a concrete floor 3.05 m below the top of the side walls. To create a water current in the flume, beneath this floor at the downstream end of the flume are two 1.7-m-diameter bow thrusters (400 hp each [1 hp = 746 W]) capable of pumping up to 14.2 m3 of filtered water per second through the test channel with the assistance of turning vanes at both ends (i.e., flume water is circulated vertically at either end). The section of flume available for testing was 24.4 m long, 6.1 m wide, and up to 2.4 m deep. In this configuration, the maximum channel velocity for the full width of the flume is 0.91 m/s. Higher flow velocities for testing the FFP turbine were achieved with temporary walls that narrowed the flume width to 2.4 m while maintaining the water depth at 2.4 m (Figure 2). With a narrower channel, the maximum current velocity that could be achieved was about 2 m/s. To minimize flow separation and turbulence, the entrance to the narrowed section had rounded walls.

Photograph of the large-flume fish testing facility as it would appear from downstream (facing into the direction of flow) showing the locations of the treatment and control injection and release points, the video cameras, and the net enclosure (insert).
The flume was equipped with a side-mounted Acoustic Doppler Current Profiler to measure current velocities and determine flow rates. Velocity measurements were recorded to verify that the flume operating conditions produced the desired current velocities with a relatively uniform distribution upstream of the test turbine. These measurements were used to develop a predicted bow thruster output curve (a rating curve that relates the flow velocity to bow thruster rpm), so that bow thruster rpm could be used to set the current velocity for each test. Once the appropriate rpm for each velocity condition was determined, a complete velocity profile was developed for each velocity condition. Velocities in the flume were measured for each test type (survival and behavioral) and velocity condition in a 3 × 3 grid to determine the average velocity profile for a given condition across the flume channel (Amaral et al. 2014). Velocity measurements were recorded with a Swoffer propeller-style velocity meter. Velocity profile measurements were recorded 6.9 m upstream of the turbine for both test types. An additional velocity profile was recorded 0.46 m in front of the turbine for the behavioral velocity conditions. The velocity measurements recorded during the survival conditions were taken with the containment net in place (see the Survival Evaluation subsection below). Additionally, velocity measurements were recorded 0.33 m upstream of the HK turbine rotor hub (center) and at a location half way between the hub and the turbine shroud that surrounds the rotor (off center) approximately 0.66 m upstream of the blades for each velocity condition. These velocity measurements were recorded in front of the turbine within the containment net enclosure to document the current velocities experienced by turbine-entrained fish during the survival trials (Figure 2).
The underwater video cameras (resolution: 640 × 480 pixels; frame rate: 29 frames/s) used to record fish movements during the survival and lighted behavioral trials were located (1) upstream of and on either side of the turbine and (2) on the flume floor downstream of the turbine. For survival testing, an additional camera was mounted inside the net enclosure at the upstream end with a partial view of the turbine blade rotational area. A dual-frequency identification sonar (DIDSON; Sound Metrics Corp., Bellevue, Washington) unit recorded the movements of fish under both light and dark test conditions at a velocity of 1.1 m/s during the behavioral trials. The DIDSON was positioned 6.9 m upstream of the turbine at a depth of 0.45 m. We also used the DIDSON during lighted trials to enable us to calculate a correction factor for entrainment counts recorded from DIDSON images under dark conditions by comparing the underwater video entrainment counts with the DIDSON counts from the lighted trials (see the next subsection).
Behavioral Testing
The behavioral trials evaluated the ability of fish to actively avoid passing through the turbine after they were released into the flume and moved downstream. In this study, fish were released from a vertical 20.3-cm-diameter tube positioned 1.45 m upstream of the FFP turbine and 40.6 cm above the centerline of the unit at the hub. The containment net was not used in these experiments, so this design allowed fish the opportunity to avoid entrainment into the HK duct (e.g., the circular cover, or shroud surrounding the turbine rotor) and through the rotor.
 (1)
(1)For the behavioral testing, three replicate trials were conducted for each set of test conditions (species, size-group [for Rainbow Trout], and current velocity; Table 1). The numbers of fish released in the trials were as follows: HSB = 50 per trial (150 per test condition), large Rainbow Trout (>200 mm FL) = 100 per trial (300 per test condition), small Rainbow Trout (≤200 mm FL) = 50 per trial (150 per test condition), and White Sturgeon = 23 per trial (69 per test condition). It was our original goal to use 100 fish per trial for all species, but our sample sizes were limited by fish availability. We obtained fish from the following sources: Osage Catfisheries, Osage Beach, Missouri (HSB); Hy-On-A-Hill Trout Farm, Plainsfield, New Hampshire (Rainbow Trout); and Professional Aquaculture Services Farm, Chico, California (White Sturgeon). We introduced fish into the flume and monitored and recorded their movements through or around the turbine with the underwater video system and/or DIDSON unit, as described previously. Each trial ended after all fish had moved downstream of the turbine, which took approximately 5 min. After completion of a trial with each species/size-group at a given current velocity, a barrier screen was lowered immediately to prevent fish from moving up or downstream of the turbine (Figures 1, 2). At the end of each trial, fish were recovered with a seine and transferred to the holding facility.
Survival Evaluation
Survival tests were conducted to estimate the fish survival, injury, and descaling rates associated with passage through the FFP turbine at two current velocities (1.5 and 2.0 m/s). Turbine passage survival was then evaluated for the three fish species 1 and 48 h poststudy. Our study evaluated the passage survival of treatment groups that were subjected to turbine entrainment (hereafter, treatment) and control groups that were not subjected to turbine entrainment (hereafter, control). In this experiment, the control group allowed for injury and mortality associated with handling and test procedures (e.g., marking, release, and collection) to be distinguished from turbine-induced injury and mortality. Initial sample sizes were determined based on an analysis of desired precision levels (±5% over 95% of the time) for survival estimates, fish costs, and availability. For Rainbow Trout tests, the goal was to use 1,000 fish for five replicate trials (100 treatment fish and 100 control fish per trial) conducted with each set of test conditions (fish size-class, approach velocity, and light condition). The target sample sizes per trial for HSB and White Sturgeon were 50 and 25, respectively, and three replicates were conducted with each species.
We marked fish with biologically inert, encapsulated photonic dyes 24 h or more prior to testing using a POW'R-Ject marking gun (New West Technology, Arcata, California). This marking system uses compressed CO2 to inject the photonic dye at the base of or into individual fins. Four dye colors and four fin location combinations were used to create 16 unique marks to distinguish treatment and control fish for each trial. Mark retention was very high; of the 4,899 treatment and control fish recovered during survival testing, only 61 (1.3%) did not have a discernible mark when recovered and were censored from the analyses. Prior to testing, we removed any fish with visible injuries or abnormal swimming behavior.
Treatment fish were released into a 2.2-cm knotless mesh containment net attached to the upstream side of the turbine through a vertical 20.3-cm-diameter release tube with an exit located 12.7 cm upstream of the turbine duct and 40.6 cm above the centerline of the unit at the hub (Figures 1, 2). The containment net prevented escape and forced downstream-moving treatment fish to pass through the turbine. Control fish were released into the flume at approximately the same time as the treatment fish. The control fish introduction system was similar to that for treatment fish, but with the exit located at the downstream end and to the side of the turbine duct.
Each group was released into the flume after the channel velocity and turbine rotational speed stabilized. Each trial was terminated after all treatment fish had passed the turbine or 10 min postintroduction, whichever came first. At the completion of each trial, fish were recovered with a seine, counted, and transferred to a holding tank where live fish were held for a 48-h observation period. Fish that died during the observation period were immediately removed from the tank.
Survival, injury, and scale loss evaluations were conducted on all recovered fish to enumerate immediate and delayed mortalities, external injuries, and percent scale loss (Table 2). Immediate mortalities were classified as any fish that died within 1 h of the completion of a test. This included any fish that were euthanized at the 1-h posttest observation due to abnormal swimming behavior (e.g., lying on tank bottom) or extensive injuries (e.g., large lacerations). We classified any fish that died between 1 and 48 h posttest as a 48-h mortality. We conducted injury and scale loss evaluations at 1 and 48 h posttest for live fish and at the time of recovery for mortalities. External injuries were recorded as bruising/hemorrhaging, lacerations, severed body, and eye damage. Using methods similar to those reported by Neitzel et al. (1985) and Basham et al. (1982), percent scale loss (<3%, 3–20%, >20–40%, and >40%) was recorded for each of three locations along the length of both sides of the body. If >20% scale loss was recorded at two or more locations, a fish was classified as descaled. During the injury evaluation, each fish was also inspected for fin mark location and color to determine its release group and test number and measured to the nearest millimeter (fork length).
| No injury | Bruising | Eye injury | Lacerations | Descaling | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Velocity (m/s) | Treatment | Control | Treatment | Control | Treatment | Control | Treatment | Control | Treatment | Control |
| Hybrid Striped Bass | ||||||||||
| 1.5 | 110 | 148 | 38 | 18 | 0 | 0 | 1 | 0 | 39 | 40 |
| 2.0 | 117 | 152 | 29 | 49 | 0 | 1 | 0 | 0 | 58 | 74 |
| Rainbow Trout | ||||||||||
| Small 1.5 | 443 | 501 | 44 | 6 | 3 | 0 | 0 | 0 | 230 | 280 |
| 2.0 | 399 | 477 | 86 | 29 | 3 | 1 | 0 | 0 | 220 | 191 |
| Large | 424 | 471 | 66 | 14 | 4 | 6 | 2 | 2 | 270 | 182 |
| 1.5 | ||||||||||
| 2.0 | 386 | 501 | 109 | 18 | 2 | 1 | 0 | 0 | 194 | 125 |
| White Sturgeon | ||||||||||
| 1.5 | 72 | 74 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 2.0 | 67 | 76 | 6 | 2 | 0 | 0 | 0 | 0 | 0 | 0 |
Data Analysis
Behavioral studies.
Our behavioral data analysis was based on both qualitative observations of fish behavior as the fish moved downstream past the turbine and quantitative estimates of entrainment and avoidance rates. Video from the underwater cameras and/or the DIDSON unit were used to count the number of fish that were entrained through the FFP turbine during each behavioral trial. Counts of entrained fish were subtracted from the total number of fish collected downstream of the turbine at the end of a test to determine the number of fish that avoided entrainment during each trial.
We used a recapture-only multistate model (Hestbeck et al. 1991; Brownie et al. 1993) in Program MARK (White and Burnham 1999) with covariates (trial, velocity, and size [the last for Rainbow Trout only]) to estimate entrainment probability (ψ), that is, the probability of moving from a nonentrained (noent) to an entrained state (ent). For instance, the probability of a fish's being entrained would be represented by ψnoent–ent. We were only interested in ψ, which was calculated from the initial release and one encounter period. We manually fixed some of the parameters in the model to 0 (e.g., ψent–noent, as it is unlikely that a fish would swim back out of the turbine once it was entrained) and others to 1 (e.g., S0 [survival at the beginning of the experiment] and ρ [the recapture probability, which was assumed to be constant throughout the study]). Because light was manipulated only in the slowest velocity, we ran separate models for each species to account for potential differences in light treatments that included only trial and size (for Rainbow Trout) as covariates. All of the differences in this study, such as those between treatment and control, flume velocities, and light conditions, were compared by means of 95% CIs, that is, we determined differences to be significant when the 95% CIs of two groups did not overlap.
Turbine passage survival.
We assessed turbine passage survival (S) using a known-fate mark–recapture model (Program MARK; White and Burnham 1999). Known-fate models allow for high-precision estimation of S probabilities and their associated confidence intervals because the status of every fish (alive or dead) can be determined with certainty at each time period. This model type allowed estimation of S both at 1 h postexperiment and over the entire 48-h study period. For this analysis, we made encounter histories using Microsoft Excel. We included the initial release (t0) plus two encounter periods (t1 and t48) and three or four covariates (trial, treatment, velocity [1.5 or 2.0 m/s], and, for Rainbow Trout only, fish size) in our encounter histories. We tested models that included no covariates, all of the covariates jointly, and each covariate separately and that allowed S to vary by time period (St) as well as remain constant in all combinations and then used a corrected Akaike's information criterion (AICc) for model selection. We chose to use AICc-based model selection to increase the accuracy of our parameter estimates and understand what factors most influenced fish injury and mortality in relation to HK turbines. The model(s) with the lowest AICc value was/were considered the best model(s), and all models with a ΔAICc value of <2 were considered to be equally parsimonious. In cases in which more than one model had a ΔAICc of <2, we obtained model-averaged parameter estimates by multiplying the estimate by the model weight for each model and summing these products. Only models with a ΔAICc of <2 were used in calculating model weights. Of the 4,899 fish that were used in this portion of the study, 62 were censored either because they could not be recovered during the trial in which they were used (N = 1) or they did not have discernable marks (N = 61, all of which were still alive at t48).
Injury and descaling attributable to turbine passage.
We used a multistate mark–recapture model with live and dead encounters (Hestbeck et al. 1991; Brownie et al. 1993) to assess the probability of fish injury (i.e., bruising, lacerations, or eye injury) during turbine passage (Program MARK; White and Burnham 1999). We estimated the transition from the uninjured state to the injured state (inj) whether fish were subjected to the turbine passage treatment (trt)or not (ctrl). We also included 4 covariates: trial, velocity (1.5 or 2.0 m/s), fork length (mm), and whether or not a fish was descaled. We included descaling as a covariate because this was a fairly common injury among the fish and it has been linked to mortality (Bouck and Smith 1979). For the purposes of this study, descaling was assessed separately from other forms of injury; thus, descaling was not captured by the injury covariate. Owing to logistical constraints with the holding tanks and the high cost associated with obtaining individual marks for such a large number of fish, individual injury data were only recorded at t1, thus leaving us with only one encounter occasion (in addition to the injury assessment prior to releasing the fish).
 (2)
(2)We used this same approach to measure the probability of descaling, that is, the probability ψuninj–descaled (from equation 2) of transitioning from a scaled state (uninj; both treatment and control fish) to a descaled state. We again included four covariates: trial, velocity, length (a continuous variable), and whether a fish was injured or not. Corrected AIC (AICc) values were used for model selection. We again made statistical comparisons between groups by examining the overlaps between 95% CIs; groups were considered to differ if there was no overlap.
Estimation of total passage survival.
 (3)
(3) (4)
(4) (5)
(5)Total passage survival, injury, and descaling probability.
 (6)
(6) (7)
(7) (8)
(8) (9)
(9)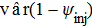 and
and 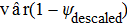 are equal to vâr(ψinj) and vâr(ψdescaled), as shown in Powell (2007).
are equal to vâr(ψinj) and vâr(ψdescaled), as shown in Powell (2007).Comparisons among hydrokinetic turbine types.
Turbine passage survival data have now been collected for fish entrained through four different pilot-scale HK turbines evaluated in a laboratory flume, two of which are ducted axial-flow designs and two of which are cross-flow units (EPRI 2011b; Castro-Santos and Haro 2015). In addition to these laboratory studies, there has also been one field study that evaluated the turbine survival of several fish species passed through a full-scale ducted axial-flow turbine (NAI 2009). The FFP, Welka UPG, and Lucid turbines evaluated in the Alden laboratory flume were all tested at approach velocities of 1.5 and 2.0 m/s, and we used these common conditions to make explicit comparisons among turbine types.
RESULTS
Behavioral Testing
Hybrid Striped Bass.
When released above the turbine, HSB actively resisted entrainment by swimming away from the turbine. Probabilities of entrainment (ψnoent–ent) were best estimated using a single model containing trial, light, and velocity as covariates (Table 3) that showed HSB having the highest probability among the tested species of being entrained through the rotor (Table 4). Model AICc values highlighted two best models of entrainment probabilities at different light levels (at the 1.1-m/s velocity; Table 4). Interestingly, the averaged estimates from these two models showed that the daytime light treatment—which also had the lowest velocities— yielded the highest entrainment probabilities (0.55), although the light and dark entrainment estimates were not significantly different (Table 4).
| Species and covariate(s) | AICc | ΔAICc | AICc weight |
|---|---|---|---|
| Behavior experiment | |||
| Hybrid Striped Bass | |||
| Trial + light + velocity | 777.29 | 0.00 | 1.00 |
| Rainbow Trout | |||
| Trial | 440.60 | 0.00 | 0.35 |
| Size + velocity | 440.93 | 0.33 | 0.21 |
| Trial + velocity | 441.29 | 0.68 | 0.18 |
| Trial + size | 442.24 | 1.64 | 0.13 |
| White Sturgeon | |||
| Trial + velocity | 159.85 | 0.00 | 0.59 |
| Trial + light + velocity | 160.54 | 0.69 | 0.41 |
| Light and behavior experiment | |||
| Hybrid Striped Bass | |||
| Trial | 396.86 | 0.00 | 0.64 |
| Trial + light | 396.86 | 1.13 | 0.36 |
| Rainbow Trout | |||
| Trial | 172.48 | 0.00 | 0.27 |
| Size | 172.57 | 0.10 | 0.25 |
| Size + light | 174.04 | 1.56 | 0.12 |
| Trial + light | 174.04 | 1.56 | 0.12 |
| Light | 174.07 | 1.59 | 0.12 |
| Trial + size | 174.16 | 1.69 | 0.11 |
| White Sturgeon | |||
| Trial | 106.40 | 0.00 | 0.43 |
| Light | 106.49 | 0.09 | 0.41 |
| Trial + light | 108.48 | 2.08 | 0.15 |
| ψnoent-ent | |||
|---|---|---|---|
| Model | Overall | Light | Dark |
| Hybrid Striped Bass | |||
| Velocity (m/s) | |||
| 1.1 | 0.59 (0.48–0.68) | 0.55 (0.33–0.71) | 0.24 (0.14–0.42) |
| 1.5 | 0.43 (0.40–0.48) | ||
| 2.0 | 0.19 (0.09–0.34) | ||
| Rainbow Trout | |||
| Size | |||
| Large | 0.01 (0.01–0.06) | 0.01 (0.01–0.03) | 0.02 (0.01–0.05) |
| Small | 0.01 (0.01–0.06) | 0.02 (0.01–0.06) | 0.02 (0.01–0.06) |
| Velocity (m/s) | |||
| 1.1 | 0.02 (0.01–0.03) | ||
| 1.5 | 0.03 (0.02–0.04) | ||
| 2.0 | 0.06 (0.04–0.08) | ||
| White Sturgeon | |||
| Velocity (m/s) | |||
| 1.1 | 0.23 (0.10–0.41) | 0.11 (0.05–0.23) | 0.14 (0.07–0.27) |
| 1.5 | 0.05 (0.03–0.09) | ||
| 2.0 | <0.01 (<0.01–0.04) | ||
Rainbow Trout.
After leaving the introduction pipe and entering the channel flow, Rainbow Trout typically exhibited positive rheotaxis and proceeded downstream in a controlled manner (head upstream and tail beating with some control of speed and directional movement). Anecdotally, of the species tested Rainbow Trout was the most active in avoiding entrainment by means of lateral and/or vertical movements. Velocity and trial were the most important covariates influencing entrainment probability and some combination of these covariates was included in all of the best models, thus suggesting that variation among trials and current velocity were important factors in whether or not a fish was entrained (Table 3). Size was included in two of the four best models and was also an important covariate influencing Rainbow Trout entrainment probability (Table 3). Interestingly, Rainbow Trout was the only species that exhibited a positive relationship between current velocity and entrainment probability. It was also the only species for which size was noted (although length was recorded for all fish in the survival trials), and although it was included in the best models there was no difference in entrainment probability between sizes (Table 4). Trial, size, and light were equally important covariates, each being found in three of the six best models examining the influence of the light treatment on Rainbow Trout entrainment (Table 3). Overall, entrainment probabilities were low, with the highest probability (0.06) being found at the 2-m/s velocity (Table 4).
White Sturgeon.
Unlike the HSB and Rainbow Trout, most White Sturgeon appeared to drift passively downstream with minimal active swimming and directional movement once entering the channel flow. White Sturgeon typically rose slightly in the water column after exiting the release pipe and followed the flow streamlines, accelerating around the outside of the turbine duct. Trial and velocity were important covariates in determining White Sturgeon entrainment probabilities and were included in both of the best models. Although it may be counterintuitive, the probability of White Sturgeon being entrained decreased with increasing velocity, and the 1.1-m/s velocity had significantly higher entrainment probabilities than either of the faster treatments (Table 4). In models that examined the impact of light on ψnoent–ent, light was included as a covariate in two of the three best models for White Sturgeon—a finding similar to those for the other species. For White Sturgeon, there was no significant difference between tests conducted under light and dark conditions (Table 4).
Survival, Injury, and Descaling Tests
Hybrid Striped Bass.
There was one best model for HSB survival, and it contained all covariates (trial, velocity, and treatment; Table 5). Compared with control fish, treatment HSB had significantly lower values of S at both t1 (treatment: 0.96 [0.91–0.98]; control: 1.00 [1.00–1.00]) and t48 (treatment: 0.91 [0.84–0.95]; control: 1.00 [1.00–1.00]). In fact, treatment HSB subjected to low-velocity conditions had significantly lower St1 estimates than treatment HSB subjected to the high velocity (slow: 0.96 [0.91–0.98]); fast: 1.00 [1.00–1.00]). The relatively low total survival estimated for HSB tested at the 1.5-m/s velocity was primarily due to the large number of delayed mortalities among treatment fish that occurred during one of the three trials. This appears to be an outlier, especially since the turbine passage survival of HSB was much higher at the higher approach velocity. We suspect that the high delayed mortality of treatment fish at the lower velocity was related to increased impingement on the enclosure net before the fish passed through the turbine and/or on the downstream barrier screen after turbine passage.
| Model | AICc | ΔAICc | AICc weight |
|---|---|---|---|
| Hybrid Striped Bass | |||
| St + trial + velocity + treatment | 233.51 | 0.00 | 1.00 |
| Rainbow Trout | |||
| St + velocity + treatment | 235.29 | 0.00 | 0.47 |
| St + size + treatment | 236.52 | 1.24 | 0.25 |
| St + trial+ velocity + treatment | 237.26 | 1.97 | 0.18 |
| White Sturgeon | |||
| St | 35.48 | 0.00 | 0.19 |
| St + trial | 35.48 | 0.00 | 0.19 |
| St + velocity | 35.48 | 0.00 | 0.19 |
| St + treatment | 35.48 | 0.00 | 0.19 |
| St + trial + treatment | 35.94 | 0.45 | 0.15 |
| St + velocity + treatment | 37.21 | 1.73 | 0.08 |
Bruising was the most prevalent injury type in treatment and control HSB (23% of all fish; Table 2), with very few incidences of eye damage or lacerations. The relatively high incidence of bruising in both treatment and control fish likely resulted from impingement on the containment net before turbine passage or on the downstream isolation screen after passage in the trial with uncharacteristically high mortality. Although this higher injury rate may be attributable to impingement at either of these two locations, it is unknown what factors contributed to excessive impingement during the one trial in which many of these injuries were observed.
The best injury models for HSB only contained trial, length, and whether a fish was descaled; velocity was not included in these models (Table 6). Model-averaged estimates of the probability of injury indicated that there were no differences between treatment and control at any length (Figure 3), and these did not change based on whether a fish was descaled or not. The probability of injury did, however, increase with fish length (Figure 4). Fish that were descaled had a much higher rate of injury than fish that were not; descaling was associated with a 0.34 (0.27–0.43) probability of visible injury in fish, compared with a 0.15 (0.11–0.20) probability of visible injury in fish with intact scales (Table 7). This high rate of injury reflected the large number of mortalities that occurred during the single trial with uncharacteristically high mortality rates (in which nearly all of the dead fish sustained bruising and/or lacerations).
| Model | AICc | ΔAICc | AICc weight |
|---|---|---|---|
| Probability of injury | |||
| Hybrid Striped Bass | |||
| Length + descaled | 767.52 | 0.00 | 0.64 |
| Trial + length + descaled | 768.69 | 1.17 | 0.36 |
| Rainbow Trout | |||
| Trial + velocity | 2,582.73 | 0.00 | 1.0 |
| White Sturgeon | |||
| Trial + velocity | 209.22 | 0.00 | 0.71 |
| Trial + velocity + length | 210.98 | 1.76 | 0.29 |
| Probability of descaling | |||
| Hybrid Striped Bass | |||
| Trial + velocity + length + injury | 6,596.95 | 0.00 | 1.00 |
| Rainbow Trout | |||
| Trial + velocity + length + injury | 5,268.06 | 0.00 | 1.00 |
| ψnoinj–inj | ||
|---|---|---|
| Model | Control | Treatment |
| Hybrid Striped Bass | ||
| Injury | ||
| Intact scales | 0.15 (0.11–0.20) | 0.15 (0.11–0.20) |
| Descaled | 0.34 (0.27–0.42) | 0.34 (0.27–0.43) |
| Overall | 0.20 (0.16–0.26) | 0.20 (0.16–0.26) |
| Rainbow Trout | ||
| Velocity | ||
| 1.5 | 0.03 (0.02–0.04) | 0.12 (0.10–0.14) |
| 2.0 | 0.05 (0.04–0.06) | 0.19 (0.17–0.22) |
| Overall | 0.04 (0.03–0.05) | 0.15 (0.14–0.17) |
| White Sturgeon | ||
| Velocity | ||
| 1.5 | <0.01 (<0.01–0.01) | 0.01 (< 0.01–0.03) |
| 2.0 | 0.04 (0.01–0.14) | 0.14 (0.06–0.28) |
| Overall | 0.01 (<0.01–0.02) | 0.03 (0.02–0.06) |

Probability of fish injury during the entrainment survival experiment with respect to fish length for control (dashed lines) and treatment (solid lines) hybrid Striped Bass, Rainbow Trout, and White Sturgeon. The shaded areas represent the 95% confidence intervals (control = dark gray, treatment = light gray) about the injury estimates. The stippled areas represent overlap between control and treatment fish, thus indicating no difference between them.

Probability of fish being descaled during the entrainment survival experiment with respect to fish length for control and treatment hybrid Striped Bass and Rainbow Trout.
The best model for estimating the probability of descaling in HSB also included all four covariates (Table 6). Both treatment and control fish had high probabilities of descaling and were more likely to be descaled at the lower velocity if they had an additional injury (Table 8). Like the injury rates, the higher descaling at the lower velocity was linked to the one trial with high mortality and injury rates. The probability of being descaled decreased as fish length increased, although there was no difference between treatment and control fish, suggesting that substantial differences in descaling were not caused by turbine passage (Figure 4).
| ψnodescale–descale | ||
|---|---|---|
| Model | Control | Treatment |
| Hybrid Striped Bass | ||
| Velocity (m/s) | ||
| 1.5 | 0.45 (0.35–0.55) | 0.39 (0.29–0.49) |
| 2.0 | 0.32 (0.24–0.42) | 0.27 (0.19–0.36) |
| Injury | ||
| Uninjured | 0.32 (0.26–0.38) | 0.26 (0.22–0.33) |
| Injury | 0.63 (0.53–0.72) | 0.57 (0.47–0.67) |
| Overall | 0.38 (0.33–0.44) | 0.33 (0.27–0.39) |
| Rainbow Trout | ||
| Velocity (m/s) | ||
| 1.5 | 0.45 (0.42–0.48) | 0.50 (0.47–0.53) |
| 2.0 | 0.38 (0.35–0.41) | 0.43 (0.40–0.46) |
| Injury | ||
| Uninjured | 0.37 (0.35–0.40) | 0.76 (0.71–0.79) |
| Injured | 0.72 (0.67–0.76) | 0.42 (0.40–0.44) |
| Overall | 0.41 (0.39–0.43) | 0.45 (0.44–0.48) |
Rainbow Trout.
There were three best survival models for Rainbow Trout; all of these contained treatment as a covariate, two contained velocity, and one each separately contained size and trial (Table 5). With the exception of small Rainbow Trout in the high-velocity treatment, which had a 48-h survival rate of 0.99 (0.97–0.99), no estimates of St1 and St48 differed from 1.00.
As with HSB, bruising was the most prevalent injury type observed for each test group, size-group, and velocity evaluated. Irrespective of species, bruising accounted for 138 of 149 (93%) recorded injuries for control fish and 382 of 398 (96%) for treatment fish (Table 2). The best model examining the probability of being injured only contained trial and velocity as covariates, suggesting that length and whether or not a Rainbow Trout was descaled were not associated with the probability of injury (Table 6; Figure 3). The numbers of Rainbow Trout recovered with visible external injuries were higher overall for treatment fish than for control fish and higher for the higher test velocity for both treatment and control fish.
The model describing the probability of being descaled that contained all four covariates was ranked as the single best model by AICc value and showed that the probability of being descaled was higher for fish with other visible injuries (Table 8). In brief, this means that while injured fish were not more likely to be descaled, descaled fish were more likely to be injured. Also, descaling probabilities were higher for treatment than control fish and for both treatment and control fish at higher velocities. Descaling probability also increased with length for both treatment and control fish (Figure 4).
White Sturgeon.
Model selection procedures identified six best models, one of which was the no-covariate, time-specific St model and three of which were single-covariate models, suggesting that covariates were unimportant to model fit for White Sturgeon survival (Table 5). The survival of White Sturgeon in our study did not differ from 1.00 under any test conditions or time period evaluated. There were no statistically significant differences in survival rates between the two test velocities. The immediate and total survival rates of the control and treatment groups were not significantly different at either current velocity.
There were two best models describing White Sturgeon injury, which included trial, velocity, and length were averaged to provide parameter estimates of injury. No descaling occurred in White Sturgeon owing to their lack of true scales, and the only injury recorded was bruising at the higher test velocity (Table 2). Whereas there were virtually no injuries at 1.5 m/s, the probabilities of injury at the higher velocity were 0.04 (0.01–0.14) for control fish and 0.14 (0.06–0.28) for treatment fish (Table 7). However, the wide confidence intervals and resulting low statistical power for the treatment ψnoinj–inj estimates limited our ability to detect significant differences between the treatment and control groups. Injuries increased with length for both treatment and control fish; however, the confidence intervals for these two groups overlapped (Figure 3), indicating no statistical difference in the estimates.
Total Passage Survival, Noninjury, and Nondescaling Probability
All species had a high probability of surviving entrainment and being uninjured following passage through the FFP turbine (Table 9). For instance, the probabilities of surviving entrainment without injury were indistinguishable from 1.00 for both Rainbow Trout and White Sturgeon. As there were no significant differences in descaling probabilities between treatment and control fish, we did not calculate total descaling probabilities. The HSB had the lowest probability of surviving entrainment (≥0.92).
| Species | Velocity | FL (mean ± SE) | Psurvival | Pnoinjury |
|---|---|---|---|---|
| Hybrid Striped Bass | 1.5 | 133.10 ± 2.68 | 0.96 (0.92–1.00) | a |
| 2.0 | 119.59 ± 2.21 | 1.00 (1.00–1.00) | a | |
| Rainbow Trout | 1.5 | 211.68 ± 1.67 | 1.00 (1.00–1.00) | 0.99 (0.99–1.00) |
| 2.0 | 208.75 ± 1.40 | 1.00 (1.00–1.00) | 0.99 (0.99–1.00) | |
| White Sturgeon | 1.5 | 125.97 ± 1.70 | 1.00 (1.00–1.00) | a |
| 2.0 | 128.38 ± 0.44 | 1.00 (1.00–1.00) | 0.99 (0.99–1.00) |
- a As there was no difference between treatment and control, the effect of turbine entrainment on Pinjury was assumed to be negligible.
Comparisons among Hydrokinetic Turbine Types
The collective data from all of the HK turbine passage survival evaluations (laboratory and field) demonstrate the effects of certain turbine design and operational features on entrainment mortality (Table 10). The FFP turbine, which has the most blades (seven) and the highest strike velocities at the blade tips, typically had the lowest survival rates, particularly in tests with Rainbow Trout (the low survival of HSB at 1.5 m/s was likely an experimental effect unrelated to turbine passage). Turbine survival was highest for the Welka UPG and Hydro Green units, which had the lowest strike velocities and fewest blades, respectively. Although there is some uncertainty in the strike velocity estimate for the Hydro Green turbine due to the assumptions made about operational parameters in the absence of actual data, high survival rates (0.99) were observed for relatively large fish (Table 10). These likely resulted from blade strike velocities that are low enough not to cause mortality (i.e., less than 5 m/s).
| Turbine | Design | Number of blades | Blade leading edge thickness (mm) | Rotor diameter (m) | Rotational speed (rpm) | Tip speed (m/s) | Tip strike speed (m/s) | Species | Fish length (mean or range; mm) | Total turbine passage survival probability |
|---|---|---|---|---|---|---|---|---|---|---|
| FFP | Ducted axial flow | 7 | 15.2 | 1.5 | 64–84 | 5.1–6.7 | 5.3–7.0 | Rainbow Trout | 170 | 0.99–1.00 |
| Rainbow Trout | 258 | 0.98–1.00 | ||||||||
| White Sturgeon | 125 | 1.00 | ||||||||
| HSB | 125 | 0.91–1.00 | ||||||||
| Welka UPG | Ducted axial flow | 4 | 12.7 | 1.5 | 15–35 | 1.2–2.8 | 1.9–3.4 | Rainbow Trout | 125 | 1.00 |
| Rainbow Trout | 239 | 0.99–1.00 | ||||||||
| LMB | 125 | 0.99–1.00 | ||||||||
| LMB | 242 | 0.99–1.00 | ||||||||
| Hydro Green | Ducted axial flow | 3 | 3.7 | 21 | 4.0 | 4.7 | YLP | 118–235 | 0.99 | |
| BLG | 115–208 | 0.99 | ||||||||
| CCF | 451–627 | 0.99 | ||||||||
| SBF | 388–482 | 0.98 | ||||||||
| BBF | 415–710 | 0.99 | ||||||||
| Lucid Spherical | Cross flow | 4 | 19.1 | 1.1 | 64–83 | 3.8–5.0 | 4.1–5.4 | Rainbow Trout | 150 | 0.99–1.00 |
| Rainbow Trout | 256 | 0.99–1.00 | ||||||||
| EnCurrent | Vertical axis, cross flow | 4 | 1.5 | 60 | ATS | 183 | 0.98a | |||
| AMS | 411 | 0.88a |
- a Not significantly different from control.
DISCUSSION
Entrainment Probability
Our study suggests that most fish will be able to escape or evade turbine entrainment, even when in close proximity to a turbine. For example, many Rainbow Trout and HSB were able to actively avoid turbine entrainment despite the relatively short distance between the release point and the turbine (1.5 m; Table 4). White Sturgeon also had high turbine avoidance rates (similar to those of Rainbow Trout), but avoidance may have been an artifact of their release position in the water column given that they did not appear to use swimming as an active turbine avoidance technique. Rather, they typically drifted upward when entering the flume flow and followed the streamlines accelerating around the FFP turbine duct without apparent directional swimming.
Behavioral tests found that turbine avoidance was similar under light and dark test conditions for all three of the species tested, demonstrating the importance of nonvisual cues in promoting positive rheotaxis in HK turbine avoidance in this study. Although vison generally provides important cues, rheotaxis is a multisensory behavior with mechanisms that vary among species as a function of the combination of the stimuli and ambient conditions (Haro et al. 1997; Bak-Coleman et al. 2013); thus, for the species we tested the acoustic noise and hydrodynamic stimuli produced by HK turbines may play roles in the positive rheotaxis responsible for avoidance, especially under low-visibility conditions. Unfortunately, the DIDSON camera could not be used effectively to observe fish during behavioral tests at 1.5 and 2.0 m/s (due to air entrainment), and we were not able to determine whether the absence of visual cues is an important factor in hindering fish from avoiding turbine entrainment at higher approach velocities.
Survival and Injury
The results of our study suggest that water velocity and fish length are important in determining whether fish become injured during or survive turbine entrainment (Tables 3, 5, 6), but because there was no significant difference between the treatment and control groups, it is not clear whether injury rates increased as a result of turbine passage (Figure 3). Still, some mortality did occur in both size-groups of Rainbow Trout at the higher velocity and velocity was included in the best subset of models (Tables 5, 6), suggesting that injury and mortality were velocity dependent in our study.
In our study, fish injuries increased with velocity, although there was no difference between the survival rates of the two size-groups of Rainbow Trout at either test velocity. The probability of injury increased with length for all three species, but only Rainbow Trout showed a significant difference between treatment and control groups. While we found no significant relationships between length and the probabilities of blade strike injury and mortality, these relationships may not have been detectable within the narrow range of fish lengths that we tested. Also, although the test flume water velocities of 1.5 and 2.0 m/s are within the range within which the FFP turbine is designed to operate (1.0–3.0 m/s), it is possible that higher fish passage mortalities would have been observed if we had been able to conduct tests at the highest FFP operating velocity.
Prior modeling studies of HK turbines have reported a positive relationship between fish length and the incidence of blade strike injury, similar to the relationship found for conventional hydropower turbines (Hecker and Allen 2005). For instance, Romero-Gomez and Richmond (2014) used a particle-tracking model to estimate the probability of blade strike injury as a function of length for a hypothetical, nonducted, 2.44-m-blade-height HK turbine. Their results suggest that greater fish lengths were associated with higher probabilities of blade strike and associated mortality (although they reported lower probabilities of strikes with increasing water velocity). There were differences in mortality and injury rates between the present study and Romero-Gomez and Richmond (2014), but differences in turbine design between the two studies (ducted in the present study versus nonducted in Romero-Gomez and Richmond 2014) preclude direct comparisons of the estimates. Moreover, the Romero-Gomez and Richmond (2014) study does not rely on empirical demonstration of the strike-to-fish-length relationship for HK turbines, so we urge caution in comparing their estimates with ours. Strike mortality has been shown to increase with the ratio of fish length to blade thickness with conventional hydropower turbines (i.e., for a given blade thickness, larger fish will have a higher probability of mortality) when strike speeds are sufficiently high to cause lethal injuries (Amaral et al. 2008; EPRI 2008, 2011c). Because the velocity vectors and hydrodynamic relationships between inflowing water and rotor blades are similar for conventional and HK turbines, we would expect a similar relationship between fish length and strike probability and mortality to hold true for HK turbines. Because validation of predictions is an essential component of robust science, testing the effects of HK turbines on a much wider range of fish lengths is a necessary next step in more fully understanding the potential impacts of these turbines on fish populations and communities.
Comparison of HK Turbine Types
The results of our study, in combination with those of past studies, suggest that fish are infrequently entrained through HK turbines and that those that are entrained pass with low incidence of injury. For example, video monitoring has indicated that benthic reef fish were able to avoid a vertical-axis, cross-flow HK turbine even when in close proximity to it (Hammar et al. 2013). In fact, no turbine blade collisions were observed by Hammar et al. (2013).
Taken together, the survival data collected during our study and prior studies indicate that the probability of survival for fish passing through a variety of HK turbine designs will be high (approaching or indistinguishable from 1.00 in many cases; Table 10) but that certain design features (e.g., the number of blades, rotational speed, and blade leading-edge thickness) can lead to slightly lower survival if they cause increases in blade strike and/or mortality from a strike. Prior flume studies have shown that fish length can influence survival rates if blade strike velocities (i.e., the speed of the turbine's blade tip) exceed 5 m/s. Even for the FFP turbine, which had a large number of blades and strike velocities at the blade tips greater than 5 m/s at the highest approach velocity tested, the estimated turbine survival probability was not distinguishable from 1.00 for the larger Rainbow Trout (mean length, 258 mm).
The turbine passage survival estimates reported for HK turbines are higher than those typically reported in field evaluations of conventional hydropower turbines (e.g., radial-flow Francis turbines and axial-flow Kaplan and other propeller-type turbines; Franke et al. 1997; EPRI 1997). We attribute the higher survival rates for HK turbines to their slower rotational speeds and strike velocities, which should lead to lower strike probabilities and mortality rates. The potential for entrained fish to be injured or killed by high shear stresses, severe turbulence, or rapid changes from high to very low pressure will be considerably less for HK turbines as well (EPRI 2011b).
CONCLUSIONS
Many of the fish in our study were able to avoid HK turbine entrainment despite features of the testing facility that may have inflated entrainment rates, such as smaller areas of safe passage around the turbine than would be the case in situ and hydrodynamic flow patterns and turbulence resulting from constrained spaces near the flume walls. On the other hand, the water velocities and blade speeds in the laboratory studies were less than those that would occur under actual deployment. A number of investigators have cautioned that migratory fish passing through an entire HK power project with large numbers of closely spaced turbines may not be able to completely avoid turbine interactions (Wilson et al. 2007; Schweizer et al. 2011; Hammar et al. 2013). Further investigations examining entrainment avoidance by more species and lengths of fish as well as the probability of escaping entrainment and subsequent injury and mortality in an array of HK turbines are warranted to more fully understand the potential impacts of these turbines on riverine and tidal fishes.
ACKNOWLEDGMENTS
We thank E. Lovelace (formerly with Free Flow Power) and T. Pham (Massachusetts Maritime Academy) for technical turbine assistance, Alabama Power for use of their DIDSON unit, and N. Lucia, J. Muise, and S. St. Jean of Alden Laboratories for laboratory technical assistance. We also thank J. Brown-Saracino, S. Bickel, and three anonymous reviewers for manuscript comments. This study was funded by the U.S. Department of Energy and Electric Power Research Institute. G.F.C., M.S.B., and B.M.P. were supported by the U.S. Department of Energy's (DOE) Energy Efficiency and Renewable Energy Office, Wind and Water Power Technologies Program through Oak Ridge National Laboratory, which is managed by UT-Battelle, LLC, for the DOE under contract DE-AC05-00OR22725. Opinions expressed are those of the authors and do not reflect those of their employers.

