

Basic characteristics of the population dynamic and state of exploitation of Moroccan white seabream Diplodus sargus cadenati (Sparidae) in the Canarian archipelago
Summary
Moroccan white seabream Diplodus sargus cadenati (n = 603) were caught off the Canary Islands from April 2000 to March 2001. Total length ranged from 46 to 404 mm. The subspecies was characterized as being dygynous with partial protandry. Overall ratio of males to females was 1 : 2.9. The reproductive season extended from December to May, with a peak in spawning activity in January–February. Fifty per cent maturity was reached at 201 mm total length in males and 216 mm in females. The length–weight relationship for all individuals was described by the following parameters: a = 0.000023, and b = 2.96, when length is given in millimeters and weight in grams. Fish of 0–12 years in age were found. The von Bertalanffy growth parameters for the entire population were: L∞ = 467 mm, k = 0.143 year−1 and t0 = −2.14 year. Growth parameters differed between males and females. For all fish, instantaneous rates of mortality were Z = 0.68 year−1, M = 0.31 ± 0.6 year−1 and F = 0.37 ± 0.6 year−1; the exploitation ratio was E = 0.54 ± 0.9. Length at first capture for all individuals was 173 mm. The stock exploited above is an assumed optimum.
Introduction
The species Diplodus sargus (Linnaeus 1758) consists of seven subspecies: D. sargus ascencionis (Val. 1830), D. sargus cadenati (de la Paz, Bauchot and Daget 1974), D. sargus capensis (Smith 1844), D. sargus helenae (Sauvage 1879), D. sargus kotschyi (Steindachner 1876), D. sargus lineatus (Val. 1830) and D. sargus sargus (L. 1758) are distributed throughout the world (Bauchot and Daget, 1971; de la Paz, 1975). Only one of these subspecies, the Moroccan white seabream D. sargus cadenati, is found off the Canary Islands (Bauchot and Hureau, 1990).
Diplodus sargus cadenati is a demersal marine fish inhabiting various types of sea bottoms, especially rock, sand and seagrass beds to depths of about 150 m, but it is more abundant in the surf zone. This subspecies is distributed along the West African coasts, from the Strait of Gibraltar to Cape Verde (absent), and around Madeira and the Canary Islands. It is also found along the European Atlantic coast, from the Bay of Biscay to the Strait of Gibraltar (Bauchot and Hureau, 1990).
Information on the biology of the D. sargus subspecies is scarce; biological research has been developed only for D. sargus sargus, D. sargus kotschyi and D. sargus capensis. No information on D. sargus cadenati has been published. All subspecies studied showed protandrous hermaproditism with a special form for sexual inversion, digynic hermaproditism, for D. sargus kotschyi and D. sargus capensis. Sexual succession takes place between 2 and 6 years of age. In all cases, size structure of the populations is unbalanced in favour of females. The species is characterized by age of first maturity between the second and the third years of life, with similar sizes between sexes in length at first maturity. Spawning takes place during the winter months (Joubert, 1981; Micale et al., 1987; Abou-Seedo et al., 1990; Martínez and Villegas, 1996; Mann and Buxton, 1998). Maximum age observed for the subspecies D. sargus sargus is 10 years in the Mediterranean (Gordoa and Molí, 1997), 12 years off the northern coast of Spain in the Cantabrian Sea (Martínez and Villegas, 1996), and 21 years for D. sargus capensis in the Indian Ocean (Mann and Buxton, 1997).
In the Canary Islands, the Moroccan white seabream is caught mainly with traps at depths ranging between 10 and 120 m. This subspecies is captured throughout the year, with significant seasonal differences in the landings. No bag limits are currently imposed on its fishery and only minimum size legislation has been implemented.
The aim of the present study was to investigate some biological aspects of D. sargus cadenati in the Canarian archipelago (Eastern Central Atlantic). Sexuality, spawning cycle, maturity, age, growth and mortality were examined in order to understand the life cycle strategy of this fish and to compare its biological aspects with the information available for other D. sargus subspecies.
Materials and methods
A total of 603 Moroccan white seabream specimens were obtained from commercial catches at different fishing ports of Gran Canaria Island between April 2000 and March 2001. Fish were caught with traps at depths of 10–104 m. For each fish, total length (TL) was measured to the nearest mm and total weight (TW) to the nearest 0.1 g. Sex and maturity stages were determined macroscopically, and gonad weight (GW) was recorded to the nearest 0.01 g. Stages of maturation were classified as follows: I, immature; II, resting; III, ripe; IV, running ripe and V, spent (Holden and Raitt, 1975). Otoliths (sagittae) were removed, cleaned and stored.
The sexual pattern was established taking into account the sex determinations. Overall sex ratio (males : females) and sex ratios by size intervals (10 mm) were determined considering the number of males and females. The spawning season was established by analysing the temporal evolution in the percentage frequency of maturity stages and values of the gonadosomatic index (GSI), calculated according to Anderson and Gutreuter (1983). For estimation of the length at first maturity (length at which 50% of the fish had become mature), a logistic function was fitted to data by means of Marquardt's algorithm for non-linear least squares parameter estimation (Saila et al., 1988).
TL and TW ranges of fish were determined. The relation of TW to TL was calculated applying the exponential regression equation (Ricker, 1973). Otoliths were immersed in an alcohol solution and read whole under a compound microscope at 11× magnification using reflected light. Age estimates were obtained by counting the number of opaque bands from the nucleus to the margin. As a rule, each otolith was read three times by the senior author; readings for a given otolith were accepted only when two readings agreed. An index of average percentage error (IAPE) was used to compare the accuracy of age determinations (Beamish and Fournier, 1981). Ageing was validated by examination of monthly changes in appearance of the edges of the otoliths. This method allowed the annual formation of the rings to be determined; once the rings were confirmed to be annual, the age of each fish was determined and assigned to a year class, taking into account the date of capture, annuli formation period, and reproductive biology of the species in the area (Morales-Nin, 1987). Thus, 1 February was considered as the birth date. The von Bertalanffy growth curve was fitted to the observed length-at-age data of the resulting age-length key by means of Marquardt's algorithm for non-linear least squares parameter estimation (Saila et al., 1988). The growth performance index (Φ) was estimated in order to compare growth parameter values obtained in the present paper with those reported for other D. sargus subspecies (Pauly and Munro, 1984).
Length frequency data of the catches were converted to age frequency using the von Bertalanffy growth parameters (Pauly, 1983). The instantaneous rate of total mortality (Z) was calculated from the length-converted catch curve using the program fisat (Gayanilo et al., 1996). Instantaneous rate of natural mortality (M) was estimated using different classical methods (Beverton and Holt, 1959; Tanaka, 1960; Taylor, 1960; Rikhter and Efanov, 1976; Pauly, 1980; Hoenig, 1983). To delineate a range of values of this rate that included its real value, the longevity for each value of the instantaneous rate of natural mortality was estimated using the method of Alagaraja (1984). The instantaneous rate of fishing mortality (F) was estimated from the difference between instantaneous rates of total and natural mortality. Length at first capture was determined from the selected ogive generated from the length-converted catch curve (Pauly, 1983).
Results
Fish ranged in size from 46 to 404 mm, and weighed between 1.4 and 1236.2 g. Males ranged between 158 and 392 mm, and between 71.9 and 877.2 g. Females ranged from 162 to 404 mm, and from 78.5 to 1236.2 g. Intersexuals ranged from 185 to 347 mm, and from 113.3 to 635.4 g.
The gonads were elongated and suspended by a dorsal mesentery in the posterior region of the visceral cavity. In general, both gonadal lobes were equally well developed.
Of the fish examined, 117 were males, 341 females and 39 intersexuals. The sex of the remaining 106 fish could not be determined macroscopically as they were immature, with very thin and translucent gonads.
The overall ratio of males to females was 1 : 2.9 and χ2 analysis showed this to be significant (χ2 = 109.5 >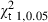 = 3.84). Males and females had the same length range; however, females predominated in all size intervals, especially in the larger ones (Fig. 1). Sex ratios by size intervals had significant departures from 1 : 1 ratio for all size categories from 150 mm, except for the 180 mm size class (
= 3.84). Males and females had the same length range; however, females predominated in all size intervals, especially in the larger ones (Fig. 1). Sex ratios by size intervals had significant departures from 1 : 1 ratio for all size categories from 150 mm, except for the 180 mm size class (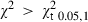 = 3.84).
= 3.84).

Number of Diplodus sargus cadenati individuals by sex and length class, and sex ratio by size intervals
Males and females with ripe gonads (stage III) were recorded between December and February, being dominant in January; ripe and running males and females (stage IV) appeared in January and became dominant in February; and spent males and females (stage V) were recorded from February to May (Fig. 2). The GSI values followed the same pattern for males and females, with values of females being higher than those of males; the highest values occurred between December and May, with a maximum in January–February (Fig. 3).

Monthly change in the maturation stages for Diplodus sargus cadenati males and females

Monthly change in the gonadosomatic index (GSI) for Diplodus sargus cadenati males and females. Vertical bars indicate standard deviation (filled bars) and maximum and minimum values
The sexual maturity ogives for males and females are shown in Fig. 4. The length at which 50% maturity is reached was 201 mm for males and at 216 mm for females. There was no significant difference in length at 50% maturity between sexes (t-test, t = 0.93 < t0.05,42 = 2.02).

Sexual maturity ogive for Diplodus sargus cadenati males and females. a, b and c are parameters, with standard error, of the logistic function of the relationship between total length (TL) and percentage of mature individuals (P): P = a/1 + [(b−a)/b] (exp−cTL)
Length–weight regression parameters for males, females and all individuals are presented in Table 1. No significant difference in the allometric coefficient was found between males and females (t-test, t = 1.76<t0.05,456 = 1.96), with females being slightly heavier than males for a given length. Negative allometric growth was observed for males, and isometric growth for females. For all fish, weight showed a negative allometric growth with length. Although the differences in the allometric coefficient were statistically significant, they were very near to isometric growth.
| a | b | SE (b) | r 2 | n | t-value | |
|---|---|---|---|---|---|---|
| Males | 0.000029 | 2.92 | 0.030 | 0.967 | 177 | 2.66* |
| Females | 0.000016 | 3.04 | 0.061 | 0.971 | 341 | 0.65 |
| All fish | 0.000023 | 2.96 | 0.019 | 0.980 | 585 | 2.10* |
- TL, total length in millimeters; TW, total weight in grams.
- *Indicates a significant deviation from isometric growth (t>t0.05,n−2>150 = 1.98).
- SE (b) is the standard error of the regression coefficient, n is the number of individuals, and t-value is the t-test estimated value.
The use of sagittal otoliths for estimating the age of the Moroccan white seabream proved straightforward, with distinct opaque and hyaline bands surrounding a white opaque nucleus. Of 603 otoliths processed for annual age estimation, 75 (12.4%) were considered unreadable, six were completely translucent, 34 were broken and 35 had poorly defined growth zones; no age estimates were obtained from these otoliths. Of the remaining 528 otoliths, 496 (93.9%) of the readings coincided at least twice, while 32 (6.1%) yielded conflicting ages and were excluded from the analysis. The index value of average percentage error was only 3.4%. Readings were equal in both right and left otoliths. Annual markings were equal in both the anterior and lateral otolith fields. Otoliths showed a seasonal variation with the formation of hyaline and opaque rings. Opaque and hyaline edges were noted in all months, but the general pattern indicated that two rings, one opaque and one hyaline, were deposited during 1 year. The opaque zone is formed between April and September, mainly in July and August, and the hyaline ring during the remaining months of the year (Fig. 5).

Monthly percentage of Diplodus sargus cadenati otoliths with opaque edges as well as monthly sea temperatures
Fish of ages 0–12 years were found. Males ranged between 2 and 11 years, and females between 2 and 12 years (Table 2). Growth parameters for males, females and all individuals are given in Table 3. Significant differences in growth parameters were found between sexes (Hotelling's T2-test, T2 = 12.2>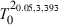 = 7.90), with females growing at a slightly faster rate than males (Fig. 6).
= 7.90), with females growing at a slightly faster rate than males (Fig. 6).
| Size (mm) | Age group (year) | T | M | F | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | ||||
| 50 | 2 | 2 | ||||||||||||||
| 60 | 3 | 3 | ||||||||||||||
| 70 | 3 | 3 | ||||||||||||||
| 80 | 3 | 3 | ||||||||||||||
| 90 | 2 | 2 | ||||||||||||||
| 100 | 2 | 2 | ||||||||||||||
| 110 | 2 | 2 | ||||||||||||||
| 120 | 2 | 1 | 3 | |||||||||||||
| 130 | 1 | 1 | 2 | |||||||||||||
| 140 | 1 | 3 | 4 | |||||||||||||
| 150 | 5 | 1 | 6 | |||||||||||||
| 160 | 23 | 3 | 26 | 3 | 1 | |||||||||||
| 170 | 22 | 7 | 29 | 4 | 3 | |||||||||||
| 180 | 16 | 17 | 1 | 34 | 9 | 8 | ||||||||||
| 190 | 5 | 41 | 3 | 49 | 11 | 31 | ||||||||||
| 200 | 2 | 43 | 6 | 51 | 11 | 33 | ||||||||||
| 210 | 1 | 27 | 11 | 39 | 7 | 27 | ||||||||||
| 220 | 21 | 22 | 43 | 11 | 27 | |||||||||||
| 230 | 12 | 39 | 2 | 53 | 14 | 33 | ||||||||||
| 240 | 2 | 26 | 3 | 31 | 9 | 19 | ||||||||||
| 250 | 13 | 7 | 20 | 5 | 14 | |||||||||||
| 260 | 3 | 15 | 18 | 4 | 12 | |||||||||||
| 270 | 19 | 1 | 20 | 5 | 13 | |||||||||||
| 280 | 7 | 2 | 9 | 2 | 7 | |||||||||||
| 290 | 4 | 3 | 7 | 2 | 4 | |||||||||||
| 300 | 2 | 5 | 1 | 8 | 2 | 5 | ||||||||||
| 310 | 4 | 2 | 6 | 2 | 4 | |||||||||||
| 320 | 1 | 5 | 6 | 3 | 3 | |||||||||||
| 330 | 2 | 2 | 4 | 1 | 3 | |||||||||||
| 340 | 1 | 3 | 4 | 2 | 2 | |||||||||||
| 350 | 1 | 1 | 2 | 1 | 1 | |||||||||||
| 360 | 2 | 2 | 4 | 1 | 3 | |||||||||||
| 370 | 1 | 2 | 3 | 3 | ||||||||||||
| 380 | 1 | 1 | 2 | 4 | 1 | 3 | ||||||||||
| 390 | 1 | 1 | 2 | 1 | 1 | |||||||||||
| 400 | 1 | 1 | 2 | 2 | ||||||||||||
| All fish | ||||||||||||||||
| n | 21 | 79 | 174 | 114 | 59 | 16 | 11 | 6 | 4 | 4 | 5 | 2 | 1 | |||
| X | 87 | 169 | 206 | 240 | 269 | 297 | 320 | 339 | 357 | 370 | 384 | 395 | 404 | |||
| SD | 19 | 14 | 18 | 21 | 15 | 13 | 11 | 8 | 11 | 9 | 8 | 7 | – | |||
| Males | ||||||||||||||||
| n | 11 | 41 | 30 | 9 | 5 | 4 | 2 | 1 | 1 | 1 | 1 | |||||
| X | 189 | 206 | 231 | 250 | 289 | 309 | 328 | 3447 | 356 | 361 | 381 | |||||
| SD | 11 | 17 | 17 | 11 | 9 | 8 | 6 | – | – | – | – | |||||
| Females | ||||||||||||||||
| n | 13 | 92 | 81 | 39 | 11 | 6 | 4 | 3 | 3 | 4 | 1 | 1 | ||||
| X | 173 | 207 | 238 | 273 | 299 | 322 | 344 | 369 | 383 | 394 | 402 | 404 | ||||
| SD | 10 | 18 | 20 | 15 | 13 | 10 | 8 | 9 | 7 | 7 | – | – | ||||
- n, the number of fish; X, the mean length by age class.
- SD, the standard deviation.
| L ∞ (mm) | k (per year) | t 0 (year) | Φ | Area | Subspecies | Authors | |
|---|---|---|---|---|---|---|---|
| Males | 447 | 0.147 | −1.89 | 4.46 | Canaries | D. sargus cadenati | Present work |
| Females | 499 | 0.129 | −2.23 | 4.41 | Canaries | D. sargus cadenati | Present work |
| All fish | 467 | 0.143 | −2.14 | 4.49 | Canaries | D. sargus cadenati | Present work |
| All fish | 309 | 0.247 | −1.04 | 4.37 | Indian Ocean | D. sargus capensis | Mann and Buxton (1997) |
| All fish | 417 | 0.245 | −0.76 | 4.62 | Catalan coast | D. sargus sargus | Gordoa and Molí (1997) |
| Males | 529 | 0.130 | −3.73 | 4.56 | Cantabrian Sea | D. sargus sargus | Martínez and Villegas (1996) |
| Females | 576 | 0.100 | −5.33 | 4.52 | Cantabrian Sea | D. sargus sargus | Martínez and Villegas (1996) |
| All fish | 485 | 0.180 | −0.58 | 4.62 | Cantabrian Sea | D. sargus sargus | Martínez and Villegas (1996) |
| All fish | 467 | 0.115 | 0.00 | 4.40 | Gulf of Lion | D. sargus sargus | Man-Wai and Quignard (1984) |
- L ∞, the asymptotic length; k, the growth rate; t0, the theoretical age at length zero and Φ the growth performance index.

von Bertalanffy growth curves for Diplodus sargus cadenati males, females and all fish. L∞, k and t0 are the parameters, with standard error, of the von Bertalanffy growth equation
The length-converted catch curve for all individuals is shown in Fig. 7. The instantaneous rate values of natural mortality obtained using different methods ranged between 0.25 and 0.76 year−1; the corresponding values of longevity were between 6 and 18 years. Some of the values of M are unrealistic because the life span expected using these data is lower than the maximum age observed for the population under natural and fishing mortalities. This is the case with the methods of Beverton and Holt (M = 0.41 years−1), and Rikhter and Efanov (M = 0.76 years−1) which result in longevity values of 11 and 6 years, respectively, lower than the age of the 12-year-old individuals sampled. The values of M obtained using the methods of Hoenig (0.37 year−1), Pauly (0.37 year−1), Taylor (0.26 year−1) and Tanaka (0.25 year−1) result in longevity values of 12, 12, 17 and 18 years, respectively, which range above the age of the oldest fish sampled. Under the effect of natural mortality only, the expected lifespan of individuals of the population studied should be higher than 12 years. For this reason, the values of M obtained using the Pauly and Hoenig methods (equal to the maximum age obtained in a population with both fishing and natural mortality) appear to underestimate the real value of M. It is thus assumed that a realistic value for the instantaneous rate of natural mortality lies in the 0.25 (lower limit) and 0.37 year−1 (upper limit) range. Fishing mortality and exploitation ratio values were estimated using a mean value for natural mortality of 0.31 year−1 as well as the limited values of M. The instantaneous rate of total and fishing mortality were, respectively, Z = 0.68 year−1 and F = 0.37 ± 0.06 year−1, and the exploitation ratio was E = 0.54 ± 0.09. Length at first capture for all individuals was 173 mm.

Length-converted catch curves for Diplodus sargus cadenati. Initial data points and the last point marked by + were not used in the regression
Discussion
The present study confirmed that Moroccan white seabream off the Canary Islands is hermaphrodite, as observed for other D. sargus subspecies in the Mediterranean, Cantabrian Sea, Red Sea and Indian Ocean (D'Ancona, 1949; Joubert, 1981; Abou-Seedo et al., 1990; Martínez and Villegas, 1996).
The presence of individuals with well-formed ovaries and residues of degenerated testes and the predominance of females at longer sizes generally confirms a protandrous development. However, the length-frequency distribution of D. sargus cadenati showed no clear separation in modal size between males and females, a diagnostic criteria for the occurrence of protandrous sex changes in fishes (Sadovy and Shapiro, 1987); both sexes attain maturity at a similar size of about 200 mm TL. This pattern has also been observed for D. sargus subspecies in the Red Sea and Indian Ocean (Joubert, 1981; Abou-Seedo et al., 1990; Mann and Buxton, 1998) and can be due to the fact that D. sargus capensis is a digynic hermaphroditic subspecies with males and females both mature from the non-functional intersexual phase, but with some males retaining the capability to change into secondary females.
The presence of males in the larger size classes confirms that sex conversion does not occur in all fish (Abou-Seedo et al., 1990). The intersexual individuals are mainly between 180 and 280 mm. Thus, sexual succession takes place mainly between 2 and 5 years of age, as described by Micale et al. (1987), Abou-Seedo et al. (1990) and Martínez and Villegas (1996) for this species. The fact that females are on average heavier than males for a given length indicates that larger size (weight) males are the first to change to females (Alekseev, 1983).
The seasonal development of the gonadosomatic index confirms winter spawning for this subspecies, as was found for others (Joubert, 1981; Man-Wai and Quignard, 1984; Abou-Seedo et al., 1990; Martínez and Villegas, 1996). The difference in index values between reproductively active males and females is similar to those observed for other sparid species off the Canaries (Pajuelo and Lorenzo, 1995a, 1996, 1998, 1999, 2000, 2001). Buxton (1990) pointed out that energy invested in gamete production by males is probably less than that invested by females. Gonadosomatic index values achieved at spawning time are very similar to those observed for moderately fecund sparid species such as Pagrus pagrus and Spondyliosoma cantharus (Manooch and Hassler, 1978; Dulcic et al., 1998), and higher than those for relatively low fecund sparid species such as Chrysoblephus puniceus and Cheimerius nufar (Garratt, 1985).
For age determination, the alternative pattern of hyaline and opaque rings was easily distinguishable on the otoliths. The opaque ring is formed when the temperature of the sea is the highest (Fig. 5) and food is most abundant (Pajuelo, 1997). The maximum age of 12 years observed for D. sargus cadenati compares well with values reported for other subspecies in the Mediterranean and in the Cantabrian Sea (Martínez and Villegas, 1996; Gordoa and Molí, 1997), however, for D. sargus capensis in South African waters a maximum of 21 years was reported (Mann and Buxton, 1997). This difference may be due to a reduction in growth at reduced temperature levels in upwelling areas, but strong upwelling events may also result in the deposition of additional rings in the otoliths, by which the ages might be overestimated. Related to this, the value of the growth performance index estimated for D. sargus of South Africa is the lowest (Table 3). In general, values of the growth performance index are very similar to those estimated with the Canarian archipelago data.
The estimated range of values of M obtained for this subspecies is very similar to that reported for other sparids off the Canary Islands with similar life history characteristics (Pajuelo and Lorenzo, 1995a, 1996, 1998, 1999, 2000). The length-converted catch curve showed a typical form and justified the estimation of a single value of Z (Pauly, 1983). The exploitation ratio depends on the assumption of the value of M. When keeping M within the assumed limits, the exploitation ratio is very near to or higher than 0.50. Gulland (1971) suggested that, as a rule of thumb, a fish stock is optimally exploited at a level of fishing mortality which generates E = 0.50, where Fopt = M. In the present study, with exception of the low limit, F > Fopt = M. More recently, Pauly (1987) proposed a lower optimum fishing mortality, Fopt = 0.4 M (E = 0.29). In this more conservative context, the range of values assumed to be realistic is higher than the value for optimal exploitation.
In the Canary Islands, a minimum size limit has been implemented for target species. In the case of the Moroccan white seabream, the minimum size regulation is of limited benefit because the minimum length, which may be legally kept, is smaller than the length at first maturity. The practical problem is that this species is only one component of the exploited multispecies demersal fish community and matures at a different size than the others (Pajuelo and Lorenzo, 1995b, 2001). Therefore, it is susceptible to exploitation at a size when many other coexisting demersal species are already mature. Measures such as a closed season or changes in fishing patterns would be desirable to safeguard the spawning stock and the recruits.
Acknowledgements
We would like to thank the people of the Fish Ecology Unit (ULPGC) who helped with sampling. This research was funded by the Universidad de Las Palmas de Gran Canaria (ULPGC). The authors are grateful to two anonymous reviewers for their critical and useful comments on the manuscript.

