

Prenatal Exposure to Perfluoroalkyl Substances Associated With Increased Susceptibility to Liver Injury in Children
Abstract
Background and Aims
Per- and polyfluoroalkyl substances (PFAS) are widespread and persistent pollutants that have been shown to have hepatotoxic effects in animal models. However, human evidence is scarce. We evaluated how prenatal exposure to PFAS associates with established serum biomarkers of liver injury and alterations in serum metabolome in children.
Approach and Results
We used data from 1,105 mothers and their children (median age, 8.2 years; interquartile range, 6.6-9.1) from the European Human Early-Life Exposome cohort (consisting of six existing population-based birth cohorts in France, Greece, Lithuania, Norway, Spain, and the United Kingdom). We measured concentrations of perfluorooctane sulfonate, perfluorooctanoate, perfluorononanoate, perfluorohexane sulfonate, and perfluoroundecanoate in maternal blood. We assessed concentrations of alanine aminotransferase, aspartate aminotransferase, and gamma-glutamyltransferase in child serum. Using Bayesian kernel machine regression, we found that higher exposure to PFAS during pregnancy was associated with higher liver enzyme levels in children. We also measured child serum metabolomics through a targeted assay and found significant perturbations in amino acid and glycerophospholipid metabolism associated with prenatal PFAS. A latent variable analysis identified a profile of children at high risk of liver injury (odds ratio, 1.56; 95% confidence interval, 1.21-1.92) that was characterized by high prenatal exposure to PFAS and increased serum levels of branched-chain amino acids (valine, leucine, and isoleucine), aromatic amino acids (tryptophan and phenylalanine), and glycerophospholipids (phosphatidylcholine [PC] aa C36:1 and Lyso-PC a C18:1).
Conclusions
Developmental exposure to PFAS can contribute to pediatric liver injury.
Abbreviations
-
- AAA
-
- aromatic amino acid
-
- ALT
-
- alanine aminotransferase
-
- AST
-
- aspartate aminotransferase
-
- BCAA
-
- branched-chain amino acid
-
- BKMR
-
- Bayesian kernel machine regression
-
- BMI
-
- body mass index
-
- CI
-
- confidence interval
-
- CV
-
- coefficient of variation
-
- GGT
-
- gamma-glutamyltransferase
-
- LC
-
- liquid chromatography
-
- LOD
-
- limit of detection
-
- MS
-
- mass spectrometry
-
- NAFLD
-
- nonalcoholic fatty liver disease
-
- OR
-
- odds ratio
-
- PC
-
- phosphatidylcholine
-
- PFAS
-
- per- and polyfluoroalkyl substances
-
- PFHxS
-
- perfluorohexane sulfonate
-
- PFNA
-
- perfluorononanoate
-
- PFOA
-
- perfluorooctanoate
-
- PFOS
-
- perfluorooctane sulfonate
-
- PFUnDA
-
- perfluoroundecanoate
Nonalcoholic fatty liver disease (NAFLD) is increasingly diagnosed at younger ages, currently affecting 8%-12% of the general pediatric population in the United States and Europe.(1) In large population studies, the prevalence of elevated serum levels of alanine aminotransferase (ALT), a biomarker of liver injury, has almost tripled among U.S. adolescents.(2) If current trends continue, NAFLD prevalence is estimated to increase by 20% over the next 10 years,(3) resulting in increased morbidity from cardiovascular disease, chronic kidney disease, and type 2 diabetes.(4) Hence, there is an urgent need to identify modifiable risk factors for liver injury and NAFLD that can be targeted for more-efficient prevention strategies.
Recently, it has been postulated that environmental chemicals with endocrine disrupting activity, commonly known as endocrine disrupting chemicals (EDCs), have the ability to promote metabolic changes that can result in fatty liver—a hypothesis referred to as the “Metabolism Disrupting Chemical” hypothesis.(5) In line with this, per- and polyfluoroalkyl substances (PFAS), a class of EDCs, concentrate considerably within the liver(6, 7) and may affect liver function.(8) PFAS are chemically and thermally stable synthetic compounds widely used in various industrial applications and consumer products, including fire-fighting foams, nonstick coatings, water- and stain-repellent textiles, and food packaging.(9) These substances have long half-lives in human tissues (estimated at up to 7 years, depending on the PFAS), and recent human biomonitoring studies show widespread exposure, with the most highly detected PFAS being perfluorooctane sulfonate (PFOS), perfluorooctanoate (PFOA), perfluorononanoate (PFNA), and perfluorohexane sulfonate (PFHxS).(10, 11)
Animal studies show that PFAS exposure causes elevated liver enzymes, liver enlargement, and hepatic steatosis,(12, 13) and that this PFAS-induced hepatotoxicity can start in utero.(14, 15) Indeed, the “Developmental Origins of Health and Disease” paradigm highlights the importance of pollutant exposures during early development in eliciting metabolic changes and increased disease risk, even after the exposure has occurred.(16) Nevertheless, in humans, evidence on the effects of PFAS on liver injury is limited to cross-sectional studies in adults(17-20) and only a few studies in children.(21, 22) Furthermore, most of these studies examined effects of each PFAS compound separately; however, humans are simultaneously exposed to several PFAS that may have additive or interactive effects.(23)
Dysregulation of lipid and amino acid metabolism has been linked to liver injury and NAFLD pathogenesis(24); however, mechanisms underlying the effects of prenatal PFAS exposure on child liver injury remain unclear. Experimental studies have shown that PFAS can increase oxidative stress while disrupting nuclear factor erythroid 2–related factor 2–regulated hepatic antioxidant defenses, exert immunotoxic effects, and induce hepatocyte caspase 3–mediated apoptosis.(25-28) Moreover, they have been shown to affect the activity of peroxisome proliferator-activated receptor-alpha and -gamma and hepatocyte nuclear factor 4-alpha, which play an important role in transcriptional regulation of lipid and amino acid metabolism.(29, 30) In a cross-sectional study of children with biopsy-proved NAFLD, we demonstrated that concurrent exposure to PFAS was associated with increased risk for nonalcoholic steatohepatitis (NASH) and higher plasma levels of phosphoethanolamine, tyrosine, phenylalanine, aspartate, and creatine.(22) Here, in a well-characterized multicenter pregnancy cohort of 1,105 mothers and their children who were followed up to the age of 6-10 years, we studied how prenatal exposure to PFAS associates with liver enzymes and alterations in serum metabolome in childhood. We hypothesized that increased prenatal plasma PFAS concentrations would be associated with increased risk of liver injury and alterations in key metabolic pathways implicated in NAFLD pathophysiology.
Participants and Methods
Study Subjects
This study was part of the Human Early-Life Exposome project (HELIX) project,(31) a collaboration across six established and ongoing longitudinal, population-based birth-cohort studies in Europe: the BiB (Born in Bradford) study in the United Kingdom, the EDEN (Étude des Déterminants pré et postnatals du développement et de la santé de l’Enfant) study in France, the INMA (INfancia y Medio Ambiente) cohort in Spain, the KANC (Kaunas cohort) in Lithuania, the MoBa (Norwegian Mother, Father and Child Cohort Study),(32) and the RHEA Mother Child Cohort study in Crete, Greece. Across these cohorts, a subcohort of 1,301 mothers and their singleton children (~200 children in each cohort) was followed in 2014-2015 for a clinical examination, a computer-assisted interview with the mother, and the collection of additional biological samples. Data collection was standardized across cohorts and performed by trained staff. The full HELIX protocol and database are described elsewhere.(31)
Our study population consisted of 1,105 (85%) mothers and their children (median age, 8.2 years; interquartile range, 6.6-9.1) from the HELIX subcohort, included based on availability of information on PFAS exposure during pregnancy and on liver enzyme levels and serum metabolomic profiling in childhood. All participating families provided written informed consent. Approval for the HELIX project was obtained from the local ethical committees at each site. Additionally, the current study was approved by the University of Southern California Institutional Review Board.
Plasma PFAS Concentrations in Pregnancy
Maternal blood samples were collected in mid-pregnancy for the INMA (mean gestational age [SD], 13.7 [2.0] weeks), MoBa (18.7 [0.9] weeks), and RHEA cohorts (14.1 [3.7] weeks) and in late pregnancy for the BiB (26.6 [1.4] weeks), EDEN (26.1 [1.2] weeks), and KANC cohorts (39.4 [1.3] weeks). Measurement of PFAS was performed at the Department of Environmental Exposure and Epidemiology at the Norwegian Institute of Public Health (NIPH) in plasma or serum samples for BiB, in serum samples for EDEN and RHEA, in whole-blood samples for KANC, and in plasma samples for MoBa. For INMA, PFAS were measured in plasma samples at the Institute for Occupational Medicine, RWTH Aachen University in Germany. Concentrations of PFOS, PFOA, PFNA, PFHxS, and perfluoroundecanoate (PFUnDA) were determined using column-switching liquid chromatography (LC) coupled to a triple-quadrupole mass spectrometer in serum or plasma and online solid-phase extraction and ultra-high-performance LC coupled with tandem mass spectrometry (MS) in whole blood.(33) A ratio of 1:1 was assumed for serum and plasma, whereas 1:2 ratios were assumed for whole-blood versus serum/plasma.(33) The limit of detection (LOD) was 0.02 μg/L for samples assessed at NIPH, whereas for samples assessed at RWTH Aachen University, LODs were 0.05 µg/L for PFNA and 0.1 µg/L for the other PFAS. We used all available PFAS measures and applied quantile regression imputation of left-censored data (QRILC) to obtain singly imputed values for samples with concentration below LOD (ranging from 0.3% to 4.7% across PFAS). QRILC imputes missing elements using random draws from a truncated distribution estimated by a quantile regression and has been shown to have high imputation accuracy.(34) Details about quality assurance and quality control have been reported(33) and are briefly described in Supporting Text S1.
Blood Sample Collection in HELIX Children
Children provided blood samples during the subcohort follow-up visit at the end of the clinical examination after a median (5th, 95th percentile) fasting time of 3.3 (2.2, 5.9) hours. Blood samples were collected and processed according to identical predefined standardized protocols across all six cohorts.
Liver Enzyme Levels in Childhood
We assessed levels of ALT, aspartate aminotransferase (AST), and gamma-glutamyltransferase (GGT) in child serum at the Biochemistry Laboratory of the Clínica Universidad de Navarra using homogenous enzymatic colorimetric methods on a Colorimetry Cobas 8000 analyzer, according to the manufacturer’s instructions (Roche Diagnostics GmbH, Mannheim, Germany). All coefficients of variation (CVs) were <3%.
Metabolite Profiling in Childhood
Child metabolite levels were quantified in serum at Imperial College of London (London, UK), using the targeted metabolomics AbsoluteIDQ p180 Kit (Biocrates Life Sciences AG, Innsbruck, Austria). The kit allows the targeted analysis of 188 metabolites of different classes, including amino acids, biogenic amines, acylcarnitines, glycerophospholipids, sphingolipids, and sum of hexoses, thus covering a wide range of analytes and metabolic pathways in one targeted assay. Of the total 188 metabolites, 42 were analyzed quantitatively by LC-electrospray ionization (ESI)-MS/MS with the use of external calibration standards at seven different concentrations and isotope-labeled internal standards for most analytes. The other 146 metabolites were analyzed by flow injection analysis-ESI-MS/MS using a 1-point internal standard calibration with 14 representative internal standards. We excluded 11 serum metabolites having a CV >30% and >30% of the data below LOD, thus leaving 177 metabolites to be used for further analysis. Median CV across these metabolites was 11.9%. Details about the assessment of serum metabolites in HELIX children have been published.(35)
Statistical Analyses
We defined liver injury risk as having any liver enzyme concentration above the 90th percentile for the study population (ALT, ≥22.7 IU/L; AST, ≥41.4 IU/L; or GGT, ≥17.1 IU/L). Maternal concentrations of PFAS were right-skewed and log2-transformed to improve model fit. Spearman’s correlation coefficients were computed to assess pair-wise correlations between individual PFAS compounds.
We examined the association of prenatal PFAS mixture exposure with liver injury risk using Bayesian kernel machine regression (BKMR), a nonparametric flexible modeling approach that can accommodate for both correlation and nonlinearity when evaluating the PFAS exposure response. Details about the BKMR models used in our study can be found in Supporting Text S2. Information on covariates was obtained through interviews, self-administered questionnaires, ad-hoc measurements, or medical records. We identified potential confounders and predictors of the outcomes of interest based on previous knowledge and a directed acyclic graph approach (Supporting Fig. S1). We included the following covariates in the models: cohort of inclusion, maternal age (in years), maternal education level (low, middle, or high), maternal prepregnancy body mass index (BMI; in kg/m2), child ethnicity (white, other), child age (in years), and child sex.
We performed several sensitivity analyses in the PFAS mixture-response association. First, we made further adjustment for child plasma levels of PFAS. Second, we additionally adjusted for child weight status (normal weight vs. overweight or obese) based on the age- and sex-specific BMI cutoffs proposed by the World Health Organization.(36) Third, we adjusted for gestational weight gain, available food indicators of maternal diet quality (consumption of fish, fruits, and vegetables; in times per week), child sedentary behavior (minutes per week), and food indicators of child diet quality (fish intake, total fruit and vegetable intakes, and total sugar-sweetened beverage consumption; in times per week). Fourth, we repeated analysis while excluding one cohort at a time to assess whether a specific cohort had a marked influence on the overall mixture effect. Fifth, we repeated analysis after stratifying by sex, given that metabolic effects in children of prenatal PFAS exposure have been previously suggested to differ by sex.(37) Sixth, we conducted stratified analyses by mid- and late-pregnancy period of PFAS assessment to assess the extent to which maternal physiological differences between these periods can affect the results.
Serum concentrations of metabolites in childhood were right-skewed and log10-transformed to improve normality. We then applied xMWAS, an automated framework for data-driven integration and differential network analysis,(38) to identify metabolites that were differentially associated with PFAS between children at high and low risk for liver injury. Details about this approach are provided in Supporting Text S3. In each group of children, we used xMWAS to first select the metabolites with the largest association to PFAS and then generate a global association network. We then compared the eigenvector centrality of each metabolite between the high- and low-risk networks to identify those with differences in their network topology. This can gauge the relative importance of a metabolite in discriminating biological processes between the two risk groups.(39, 40) If a metabolite was not present in one of the networks, its eigenvector centrality was considered 0. Metabolites with an absolute value of difference in their eigenvector centrality >0.2 were considered to have differential contribution to, and thus disparate importance in, the high-risk group as compared to low risk.(39) Upon identification of the differentiating metabolites in the high- versus low-risk group, we annotated them to their Kyoto Encyclopedia of Genes and Genomes (KEGG) and Chemical Entities of Biological Interest identifiers and performed pathway overpresentation analysis, using ConsensusPathDB,(41) to characterize dysregulated metabolic pathways that are both associated with PFAS exposure and risk for liver injury. KEGG pathways with more than two annotated metabolites and a P value < 0.05 based on the hypergeometric test were kept for further evaluation.
Last, to provide insight into the joint contribution of prenatal PFAS and the metabolites to liver injury risk, we performed an integrated latent variable analysis to estimate latent unknown clusters(42) of children associated with increased susceptibility to liver injury by incorporating information on their prenatal PFAS mixture exposure and the metabolites identified from the network analysis. Details about this method are provided in Supporting Text S4.
All analyses were done using R software (version 3.5.3; R Foundation for Statistical Computing, Vienna, Austria). We used the software packages bkmr for BKMR analysis, xMWAS for network analysis, and LUCIDus for the integrated latent variable analysis.
Results
Study Population Characteristics
No differences in liver enzyme levels were observed between boys and girls. A total of 253 children (22.8%) were classified as being at risk for liver injury. The percentage of children at risk for liver injury was highest in the Greek cohort (31.6%) and lowest in the Lithuanian cohort (4.4%; Table 1). Children at risk for liver injury were more likely to be nonwhites, overweight or obese, and born from mothers with low educational status and higher BMI prepregnancy.
| All (N = 1,105) | Liver Injury Risk* | |||
|---|---|---|---|---|
| Low (n = 852) | High (n = 253) | P for Difference† | ||
| Cohort of inclusion | <0.001 | |||
| BiB, UK | 95 (8.6) | 40 (4.7) | 55 (21.7) | |
| EDEN, France | 196 (17.7) | 154 (18.1) | 42 (16.6) | |
| INMA, Spain | 212 (19.2) | 164 (19.3) | 48 (19) | |
| KANC, Lithuania | 166 (15) | 155 (18.2) | 11 (4.4) | |
| MoBa, Norway | 271 (24.5) | 254 (29.8) | 17 (6.7) | |
| RHEA, Greece | 165 (14.9) | 85 (10) | 80 (31.6) | |
| Maternal characteristics | ||||
| Age at birth (years) | 31.0 (4.7) | 31.1 (4.7) | 30.0 (4.8) | 0.93 |
| Prepregnancy BMI (kg/m2) | 24.5 (4.8) | 24.1 (4.4) | 25.8 (5.7) | <0.001 |
| Normal weight (<25 kg/m2) | 703 (63.6) | 564 (66.2) | 139 (54.9) | |
| Overweight (≥25-<30 kg/m2) | 262 (23.7) | 198 (23.2) | 64 (25.3) | |
| Obesity (≥30 kg/m2) | 140 (12.7) | 90 (10.6) | 50 (19.8) | |
| Maternal smoking in pregnancy | ||||
| No | 948 (85.8) | 738 (86.6) | 210 (83.0) | 0.15 |
| Yes | 157 (14.2) | 114 (13.4) | 43 (17.0) | |
| Education | <0.001 | |||
| Low | 124 (11.2) | 83 (9.7) | 41 (16.2) | |
| Medium | 382 (34.6) | 277 (32.5) | 105 (41.5) | |
| High | 599 (54.2) | 492 (57.8) | 107 (42.3) | |
| Parity | 0.11 | |||
| Nulliparous | 509 (46.0) | 398 (46.7) | 112 (44.3) | |
| Multiparous | 596 (54.0) | 454 (53.3) | 141 (55.7) | |
| Child characteristics | <0.001 | |||
| Age (years) | 8.2 (1.6) | 8.3 (1.6) | 7.9 (1.7) | |
| Sex | 0.45 | |||
| Male | 597 (54) | 455 (53.4) | 142 (56.1) | |
| Female | 508 (46) | 397 (46.6) | 111 (43.9) | |
| Race/ethnicity | <0.001 | |||
| White | 1,031 (93.3) | 817 (95.9) | 214 (84.6) | |
| Other | 74 (6.7) | 35 (4.1) | 39 (15.4) | |
| BMI (kg/m2) | 17 (2.6) | 16.8 (2.2) | 17.9 (3.5) | <0.001 |
| Weight status‡ | <0.001 | |||
| Normal weight | 878 (79.5) | 712 (83.6) | 166 (65.6) | |
| Overweight | 161 (14.6) | 112 (13.2) | 49 (19.4) | |
| Obesity | 66 (6.0) | 28 (3.3) | 38 (15.0) | |
| Liver enzyme concentrations | ||||
| ALT (IU/L) | 15.7 (6.2) | 13.8 (3.6) | 22.2 (8.5) | <0.001 |
| AST (IU/L) | 30.8 (9.3) | 28.2 (5.1) | 39.7 (13.6) | <0.001 |
| GGT (IU/L) | 12.7 (3.6) | 11.7 (2.2) | 16.2 (5.1) | <0.001 |
- Data are expressed as mean (SD) or n (%).
- * Liver injury risk was defined as any liver enzyme level above the 90th percentile.
- † P value for difference between the low- and high-liver-injury risk groups was calculated using the chi-square test for categorical characteristics and the Mann-Whitney U test for continuous ones.
- ‡ Weight status was defined according to the World Health Organization BMI cutoffs.
Maternal blood PFAS concentrations of the study population are displayed in Supporting Table S1. Median (25th, 75th percentile) concentrations of PFOA, PFOS, PFHxS, PFNA, and PFUnDA in pregnancy were 2.38 (1.45, 3.45), 6.74 (4.43, 10.35), 0.59 (0.34, 0.93), 0.72 (0.47, 1.11), and 0.20 (0.13, 0.30) ng/mL, respectively. Pair-wise correlations between the PFAS compounds revealed moderate-to-high correlations, with PFOS-PFHxS and PFOA-PFHxS correlations being the highest (Spearman’s rho = 0.70 and 0.65, respectively; Supporting Table S1).
Prenatal PFAS Exposure and Liver Injury Risk in Children
Higher prenatal exposure to PFAS mixture was associated with increased risk for liver injury during childhood (Fig. 1). In Fig. 1A, we display the estimated change in the risk as maternal concentrations of all five PFASs were simultaneously fixed at the same percentile (over a range between 0.3 and 0.75, with 0.05% increments) and compared to when those PFAS concentrations were each at their 25th percentile. PFNA and PFOA had the greatest contribution to the mixture effect, as suggested by their posterior inclusion probabilities in the mixture-response function for liver injury risk (Fig. 1A). Children who were highly exposed to prenatal PFAS mixture (75th percentile) had 2-fold higher risk for liver injury (odds ratio [OR], 2.0; 95% confidence interval [CI], 1.53-2.61) compared to children with low levels of exposure (25th percentile). Similar positive associations with the prenatal PFAS mixture exposure were found when we analyzed the three liver enzymes as separate outcomes (Fig. 1B-D). When exploring the effect of each of the separate PFAS while holding the remaining PFAS within the mixture constant, increased risks were also observed for PFNA and PFOA (Supporting Fig. S2A). In further analyses examining synergism between PFAS within the mixture, we observed that the effects of PFHxS, PFOA, PFOS, and PFUnDA were more pronounced at higher levels of PFNA (Supporting Fig. S2B).


Correlations between maternal and child blood PFAS were low to moderate, with the highest correlations being observed for maternal PFOS with child PFHxS and PFOS (Spearman’s rho = 0.53 and 0.50, respectively). Effect estimates for the prenatal PFAS mixture on liver injury risk did not materially change when we adjusted for child PFAS levels (Supporting Fig. S3A). Effect estimates also did not materially change when we took into account weight status in childhood (Supporting Fig. S3B). Additional adjustment for gestational weight gain, maternal diet-quality indicators, and child lifestyle risk factors for NAFLD, including sedentary behavior and diet-quality indicators, did not change considerably the results (Supporting Fig. S3C). Significant associations of prenatal PFAS mixture exposure with increased liver injury risk were also observed when we repeated the analysis excluding one cohort at a time, as well as when we excluded the two cohorts contributing most to the cases of increased liver enzymes (Supporting Fig. S3D). In stratified analysis by sex, we found that the effect of prenatal PFAS mixture was greater in girls than in boys, although effect estimates were in the same direction and had overlapping CIs (Supporting Fig. S3E). When we stratified by trimester of pregnancy for PFAS assessment, we observed that effect estimates were slightly stronger when maternal PFAS concentrations were assessed in mid-pregnancy (first to second trimester) compared to late pregnancy, but estimates had the same direction and overlapping CIs (Supporting Fig. S3F).

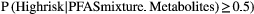 compared to those with low probability of inclusion. Median μmol/L values (25th-75th percentile) for children with low versus high inclusion probability were 223.0 (195.0, 265.0) versus 264.0 (220.0, 320.3) for valine, 127.0 (109.0, 155.0) versus 157.0 (126.0, 201.5) for leucine, 70.8 (59.94, 89.4) versus 90.6 (70.2, 115.3) for isoleucine, 67.4 (58.6, 76.3) versus 77.1 (67.5, 88.7) for tryptophan, 63.1 (54.8, 72.0) versus 72.1 (63.4, 85.9) for phenylalanine, 0.52 (0.35, 0.77) versus 0.81 (0.50, 1.71) for acetylornithine, 57.8 (49.7, 69.2) versus 67.8 (57.5, 81.7) for PC aa C36:1, and 16.6 (13.6, 20.3) versus 19.3 (16.4, 22.9) for Lyso-PC a C18:1.
compared to those with low probability of inclusion. Median μmol/L values (25th-75th percentile) for children with low versus high inclusion probability were 223.0 (195.0, 265.0) versus 264.0 (220.0, 320.3) for valine, 127.0 (109.0, 155.0) versus 157.0 (126.0, 201.5) for leucine, 70.8 (59.94, 89.4) versus 90.6 (70.2, 115.3) for isoleucine, 67.4 (58.6, 76.3) versus 77.1 (67.5, 88.7) for tryptophan, 63.1 (54.8, 72.0) versus 72.1 (63.4, 85.9) for phenylalanine, 0.52 (0.35, 0.77) versus 0.81 (0.50, 1.71) for acetylornithine, 57.8 (49.7, 69.2) versus 67.8 (57.5, 81.7) for PC aa C36:1, and 16.6 (13.6, 20.3) versus 19.3 (16.4, 22.9) for Lyso-PC a C18:1.Prenatal PFAS Exposure and the Child Serum Metabolome
We observed that the network of children at high risk for liver injury involved 66 metabolites that were associated with PFAS (Fig. 2A; Supporting Table S2). In comparison, the network of low risk involved 34 metabolites (Fig. 2B; Supporting Table S3). A total of 29 metabolites had the greatest change in eigenvector centrality across the two networks (|delta eigenvector centrality| ≥0.2). These metabolites included five amino acids, one biogenic amine, 19 glycerophospholipids, three sphingomyelins, and one hexose and exhibited both positive and negative associations with maternal serum PFAS in children at high risk for liver injury (Fig. 2C).
Pathway analysis of these metabolites showed that the most dysregulated pathways were related to protein and amino acid metabolism (Fig. 2D). Specifically, six pathways, including protein digestion and absorption, aminoacyl-tRNA (transfer RNA) biosynthesis, valine, leucine, and isoleucine biosynthesis, valine, leucine, and isoleucine degradation, and phenylamine, tyrosine, and tryptophan biosynthesis, were over-represented in the network of high liver injury risk. PFAS-associated metabolic perturbations were also observed for some lipid metabolism pathways, including glycerophospholipid metabolism, ether lipid metabolism, and sphingolipid signaling pathway.
Identification of Children at Risk for Liver Injury Based on Prenatal PFAS Exposure and Serum Metabolome
The integrated latent variable analysis estimated two subgroups of children (Fig. 3A). The high-risk subgroup had 56% increased risk for liver injury (OR, 1.56; 95% CI, 1.21-1.92) compared to the low-risk subgroup. This group was characterized by increased prenatal PFAS levels and increased child serum levels of branched-chain amino acids (BCAAs; valine, leucine, and isoleucine), the aromatic amino acids (AAAs) tryptophan and phenylalanine, biogenic amine acetylornithine, and glycerophospholipids phosphatidylcholine (PC) aa C36:1 and Lyso-PC a C18:1 (Fig. 3B; Supporting Table S4).
Discussion
In this well-characterized multicenter cohort of European mothers and their children, we demonstrate that maternal exposure to PFAS mixture during pregnancy is associated with increased liver injury risk in childhood. We also integrated maternal PFAS blood concentrations with targeted serum metabolomic profiling in children and show that prenatal PFAS exposure is associated with alterations in key amino acids and lipid pathways characterizing liver injury risk. Given that the prevalence of NAFLD in children is rapidly increasing(1) and that PFAS can efficiently cross the placenta barrier and deposit to fetal tissues,(6) these results have potential implications for public health and prevention policy.
PFAS have been in production for over 55 years, resulting in ubiquitous exposure levels. Maternal blood concentrations of PFAS in our study population were lower than those reported in the U.S. National Health and Nutrition Examination Survey (NHANES) female population over the period 1999-2010, when the recruitment of participating mothers in our study occurred, and slightly higher than recent U.S. levels (collection period 2011-2016).(11) The in utero or early-life period is a critical developmental period of high vulnerability to PFAS. In line with our findings, animal studies have shown that prenatal exposure to PFOA and PFOS causes hepatocellular injury and fatty liver in the offspring.(14, 15) Previous human cross-sectional studies have also shown positive associations of plasma or serum concentration of PFAS, including PFOS, PFOA, and PFNA, with liver enzymes levels in U.S adolescents and adults in NHANES,(19, 43) adult populations from Sweden,(20) and U.S communities drinking PFAS-contaminated water.(17, 18) In a previous U.S. study involving children with physician-diagnosed NAFLD, we showed that plasma PFOS and PFHxS levels were associated with increased risk for NASH.(22) The only pregnancy cohort study conducted previously found that maternal serum PFAS concentrations were not associated with ALT levels in U.S children.(21) A major strength of the present study is that the complicated interplay of PFAS exposure was treated in the analysis as a mixture, compared with previous studies that studied them only as single independent compounds. This approach is appropriate, given that in the real world, PFAS occur together as contaminant mixtures in soil, water, and consumer products, are highly correlated, and, as observed in our study, they may exert synergistic hepatotoxic effects.
We found a slightly stronger association of prenatal PFAS mixture in girls than in boys, findings consistent with previous research showing that females are more susceptible than males to PFAS hepatotoxicity.(44) The liver is a sexually dimorphic organ, given that it expresses both androgen and estrogen receptors and is responsive to sex steroids. Experimental studies have shown that PFAS interfere with sex steroid pathways by disturbing expression of genes involved in sex steroid biosynthesis and activity of sex steroid receptors,(30, 45) which, in turn, could confer sex differences in PFAS hepatotoxicity.(46)
The exact mechanisms through which PFAS can affect liver injury and NAFLD remain unclear. Among children with NAFLD, we previously showed that plasma PFAS were associated with alterations in several amino acid pathways (e.g., valine, leucine and isoleucine degradation, and tyrosine metabolism) and glycerophospholipid metabolism.(22) Our study confirms these results, showing that not only childhood exposure, but also prenatal exposure to PFAS are associated with alterations in amino acid (e.g., valine, leucine, and isoleucine degradation and biosynthesis; phenylalanine, tyrosine, and tryptophan biosynthesis) and lipid (e.g., glycerophospholipid) metabolism. These are key metabolic pathways that have been well linked to liver injury and NAFLD pathogenesis in both human and animal studies.(24) The integrated latent analysis identified a high-risk profile of children characterized by high prenatal PFAS exposure and increased child serum concentrations of BCAAs, AAAs, and glycerophospholipids. These results strengthen an emerging body of evidence from animal(47) and human studies,(22, 48) showing that PFAS exposure is associated with alterations in BCAA, AAA, and glycerophospholipid levels. Circulating levels of BCAAs and AAAs have been reported to be elevated in children with NAFLD.(49, 50) BCAAs and AAAs are associated with hepatic insulin resistance, a key pathophysiological feature of NAFLD, possibly as a result of impaired hepatic tricarboxylic acid cycle metabolism and mitochondrial function.(51, 52) PFAS disrupt hepatic lipid metabolism by interacting with peroxisome proliferator-activated receptors and other receptors attributable to their structural similarities with fatty acids.(8) Moreover, abnormal glycerophospholipid levels can induce hepatic lipotoxicity and inflammation, the hallmark of NAFLD, through endoplasmic reticulum stress activation and inflammatory extracellular vesicle release.(53) Consequently, altered glycerophospholipid metabolism has been reported in both blood and liver tissue samples from NAFLD patients.(50, 54)
Strengths of our study include the multicentric, prospective design involving mother-child pairs of six countries spanning north to south in Europe, with comprehensive assessment of PFAS exposure in a critical developmental time period (pregnancy) and detailed information regarding covariates and predictors of the outcome during follow-up. When we adjusted for childhood exposure to PFAS, results did not change, underlying the importance of the pregnancy as a key time period of exposure for the development of liver injury. The integrated analyses of chemical exposures and metabolomics, with the goal of identifying latent variables representing distinct groups of children at risk for liver injury, have the potential to offer a personalized paradigm with potential clinical application to the identification of pediatric populations at risk for liver injury.
Although large in scale, our study has a number of limitations. As in any observational study, residual confounding cannot be ruled out, and it might have biased the observed associations. However, our results remained similar following adjustment for a large variety of social and lifestyle factors and are in accordance with previous experimental and human evidence on PFAS hepatotoxicity; this argues against residual confounding as the sole explanation for our results. We characterized liver injury based on serum liver enzymes levels given that the current diagnostic gold-standard liver biopsy for NAFLD has well-known limitations of high cost, risk, and ethical restrictions in large population studies, such as HELIX. However, we do not expect any misclassification in the outcome to be informed by prenatal PFAS status. Although we used a longitudinal study design, metabolomic biomarkers in children were assessed at the same time point as liver enzyme levels, thus limiting the ability to disentangle any mediating effect of the observed PFAS-associated metabolic perturbations on liver injury. We acquired the serum metabolomics data using a standardized, targeted LC-MS/MS assay, which exhibits high sensitivity and specificity of the quantification, provides explicit metabolite identification, and has high interlaboratory reproducibility.(55) Although this analytical method has been widely used in large-scale epidemiology studies, thereby facilitating future comparisons, it is limited in the number of metabolites identified and provided partial coverage of the metabolome. Supplementing the current study with untargeted metabolomic approaches, such as high-resolution MS, in the future would help expand metabolite coverage and capture more pathways potentially relevant to PFAS hepatotoxicity.
In conclusion, our results suggest that in utero exposure to PFAS can contribute to liver injury in childhood and that alterations in BCAAs, AAAs, and glycerophospholipids may jointly characterize this association. The study addresses a critical gap in our current understanding of the etiology of the NAFLD epidemic in youth and has potential to lead to avenues for fatty liver disease prevention and treatment starting early in life.
Acknowledgment
We acknowledge the input of the entire HELIX consortium. We are grateful to all the participating families in the six cohorts (BiB, EDEN, INMA, KANC, MoBa, and RHEA cohorts), that took part in this study. We are equally grateful to all the fieldworkers for their dedication and efficiency in this study. A full roster of the INMA and RHEA study investigators can be found at http://www.proyectoinma.org/en/inma-project/inma-project-researchers/ and http://www.rhea.gr/en/about-rhea/the-rheateam/, respectively. The Born in Bradford study is only possible because of the enthusiasm and commitment of the participating children and parents. We are grateful to all the participants, health professionals, and researchers who have made Born in Bradford happen. We are also grateful to all the participating families in Norway who take part in the ongoing MoBa cohort study. We are grateful to all the participants, health professionals, and researchers who have participated in the Kaunas KANC cohort. We thank all the children and families participating in the EDEN-HELIX mother-child cohort. We are grateful to Joane Quentin, Lise Giorgis-Allemand, and Rémy Slama (EDEN study group) for their work on the HELIX project. We thank Sonia Brishoual, Angelique Serre, and Michele Grosdenier (Poitiers Biobank, CRB BB-0033-00068, Poitiers, France) for biological sample management and Prof. Frederic Millot (principal investigator), Elodie Migault, Manuela Boue, and Sandy Bertin (Clinical Investigation Center, Inserm CIC1402, CHU de Poitiers, Poitiers, France) for planning and investigational actions. We are also grateful to Veronique Ferrand-Rigalleau, Celine Leger, and Noella Gorry (CHU de Poitiers, Poitiers, France) for administrative assistance. We also acknowledge the commitment of the members of the EDEN Mother-Child Cohort Study Group: I. Annesi-Maesano, J.Y. Bernard, J. Botton, M.A. Charles, P. Dargent-Molina, B. de Lauzon-Guillain, P. Ducimetière, M. de Agostini, B. Foliguet, A. Forhan, X. Fritel, A. Germa, V. Goua, R. Hankard, M. Kaminski, B. Larroque, N. Lelong, J. Lepeule, G. Magnin, L. Marchand, C. Nabet, F. Pierre, M.J. Saurel-Cubizolles, M. Schweitzer, and O. Thiebaugeorges.
Author Contributions
L.C. and R.M. conceived the concept. N.S., D.V.C., B.K., and L.C. designed analytical and statistical methods. N.S. and K.M. analyzed the data, and D.V.C. supervised the statistical analysis. N.S. and L.C. wrote the first draft of the paper. M.V. coordinated the HELIX project. M.V., O.R., and L.M. coordinated the HELIX data collection. H.K. and A.P.S. generated and reviewed the primary metabolomics data. All authors contributed to data collection and/or interpretation of the results. All authors contributed to and approved the manuscript. N.S. and L.C. had full access to all the data in the study and had final responsibility for the decision to submit for publication.

