

Metastable neural dynamics underlies cognitive performance across multiple behavioural paradigms
Funding information: Department for Employment and Learning, Northern Ireland; National Institute of Mental Health, Grant/Award Number: MH080838; Science Foundation Ireland, Grant/Award Number: 11/RFP.1/NES/3194
Abstract
Despite resting state networks being associated with a variety of cognitive abilities, it remains unclear how these local areas act in concert to express particular cognitive operations. Theoretical and empirical accounts indicate that large-scale resting state networks reconcile dual tendencies towards integration and segregation by operating in a metastable regime of their coordination dynamics. Metastability may confer important behavioural qualities by binding distributed local areas into large-scale neurocognitive networks. We tested this hypothesis by analysing fMRI data in a large cohort of healthy individuals (N = 566) and comparing the metastability of the brain's large-scale resting network architecture at rest and during the performance of several tasks. Metastability was estimated using a well-defined collective variable capturing the level of 'phase-locking' between large-scale networks over time. Task-based reasoning was principally characterised by high metastability in cognitive control networks and low metastability in sensory processing areas. Although metastability between resting state networks increased during task performance, cognitive ability was more closely linked to spontaneous activity. High metastability in the intrinsic connectivity of cognitive control networks was linked to novel problem solving or fluid intelligence, but was less important in tasks relying on previous experience or crystallised intelligence. Crucially, subjects with resting architectures similar or 'pre-configured' to a task-general arrangement demonstrated superior cognitive performance. Taken together, our findings support a key linkage between the spontaneous metastability of large-scale networks in the cerebral cortex and cognition.
1 INTRODUCTION
The brains of subjects at 'cognitive rest' display circumscribed patterns of neural activity or resting state networks (Fox et al., 2005; Fox & Raichle, 2007) that broadly overlap with task-based activations (Cole, Bassett, Power, Braver, & Petersen, 2014; Smith et al., 2009). Somehow these large-scale networks of the brain rearrange themselves on a fixed anatomical structure to support internal processes relevant to cognition (Bola & Sabel, 2015; Braun et al., 2015; Cohen, 2018; Cohen & D'Esposito, 2016; Lewis, Baldassarre, Committeri, Romani, & Corbetta, 2009; Sadaghiani & Kleinschmidt, 2013; Spadone et al., 2015). One proposal is that neuronal assemblies are dynamically bound into coherent coordinative structures known as neurocognitive networks (Bressler & Kelso, 2001, 2016). The concept of the neurocognitive network represents an important compromise between two antagonistic viewpoints: the first, localisation, which holds that complex cognitive functions are localised to specific regions of the brain, the second, globalism, which posits that complex functions are distributed and arise through global coordination (Bressler & Mcintosh, 2007; McIntosh, 1999, 2000, 2004, 2007). Neurocognitive networks realise a type of dynamics where the brain's tendencies towards integration and segregation are simultaneously realised. Local areas are permitted to express their intrinsic functionality yet also couple together and coordinate globally. Cognition, in this context, is the real-time expression of distributed local areas whose states of mutual coordination are adjusted dynamically over time (Bressler & Tognoli, 2006). An important challenge is to understand how these local areas become dynamically linked in the execution of particular cognitive operations, and equally, how these patterns of dynamic connectivity evolve over time (Cabral, Kringelbach, & Deco, 2017; Gonzalez-Castillo & Bandettini, 2017).
The coordination of neurocognitive networks appears to arise from a dynamic regime that balances counteracting tendencies towards integration and segregation (Shanahan, 2010; Sporns, 2013; Tognoli & Kelso, 2014a; Tononi, Edelman, & Sporns, 1998; Tononi, Sporns, & Edelman, 1994). Empirical and theoretical accounts indicate that the brain derives this behaviour from its identity as a complex dynamical system operating in the metastable regime of its coordination dynamics (Kelso, 1995, 2012; Kelso & Tognoli, 2007; Shine, Koyejo, & Poldrack, 2016; Tognoli & Kelso, 2009; Tognoli & Kelso, 2014b; Shine et al., 2016b). The concept of metastability represents an important theoretical solution to the requirement that local areas operate independently yet also combine and behave synergistically (Kelso, 2012; Kelso & Tognoli, 2007; Tognoli & Kelso, 2014b). Metastability is also important as an observable phenomenon, furnishing a dynamical explanation for how large-scale brain regions coordinate their activity in space and time to support cortical function (Bressler & Kelso, 2001, 2016; Jirsa & McIntosh, 2007; Kelso, 2008; Tognoli & Kelso, 2009). In the language of coordination dynamics, metastability refers to a coupled or collective oscillatory activity which falls outside its equilibrium state for dwell times that depend on distance from equilibrium (Kelso, 1995). The overall dynamic stability~flexibility of these systems, where the tilde symbolizes the dynamic nature of this complementary pair, may be estimated by calculating a well-defined collective variable or order parameter (Cabral, Hugues, Sporns, & Deco, 2011; Kuramoto, 1984; Shanahan, 2010; Wildie & Shanahan, 2012). The Kuramoto order parameter captures the average phase of a group of oscillators to quantify how 'phase-locked' they are at a given moment in time. Accordingly, the variation in this order parameter has been proposed as a measure of a system's metastability and the mean of the phase-locking across time as a measure of the system's overall synchrony. Metastability is high in a system that visits a range of different states over time (dynamic flexibility) whereas both highly ordered and highly disordered states are characterised by low metastability (dynamic stability), and high and low phase synchrony, respectively. These mutually related dynamics admit an interpretation at both the functional level, in terms of neural flexibility, and at the cognitive level, in terms of behavioural flexibility. A concrete example of a metastable dynamical system is the 'winnerless competition' (Rabinovich, Huerta, Varona, & Afraimovich, 2008; Rabinovich, Varona, Selverston, & Abarbanel, 2006). However, metastable phenomena may emerge from a variety of underlying mechanisms (where certain conditions are satisfied) and it is in this broader sense that we use the term (Balaguer-Ballester, Moreno-Bote, Deco, & Durstewitz, 2018; Deco & Jirsa, 2012; Deco & Kringelbach, 2016; Friston, 1997; Kringelbach, McIntosh, Ritter, Jirsa, & Deco, 2015; Stratton & Wiles, 2015).
The concept of phase synchronisation was originally introduced in physics to study the behaviour of weakly coupled oscillators (Rosenblum, Pikovsky, & Kurths, 1996). The original motivation was to compare the temporal structure of two time series by removing information related to amplitude (Varela, Lachaux, Rodriguez, & Martinerie, 2001). Signal processing techniques such as the Hilbert transform make it possible to separate a time series into its amplitude and phase by converting the real signal into its complex analytic version (Boashash, 1992). Importantly, unlike correlation-based sliding-window analysis, which mandates an arbitrary choice of window length, the phase synchronisation approach provides time-resolved functional connectivity at the same resolution as the input narrowband fMRI signal (Glerean, Salmi, Lahnakoski, Jaaskelainen, & Sams, 2012). Moreover, unlike correlation, which is a linear measure of association between variables, phase synchronisation is a measure of statistical dependence that is sensitive to both linear and non-linear relationships (Pereda, Quiroga, & Bhattacharya, 2005). Recently, the phase synchronisation approach has successfully identified changes in the time-varying properties of brain connectivity associated with several neural disorders (Alderson, Bokde, Kelso, Maguire, & Coyle, 2018; Córdova-Palomera et al., 2017; Demirtaş et al., 2017; Hellyer, Scott, Shanahan, Sharp, & Leech, 2015; Koutsoukos & Angelopoulos, 2018; Lee, Doucet, Leibu, & Frangou, 2018).
Theoretical accounts stipulate that metastability at rest corresponds to an optimal exploration of the dynamical repertoire inherent in the static structural linkages of the anatomy where the probability of network switching is maximal (Cabral et al., 2011; Deco, Kringelbach, Jirsa, & Ritter, 2017; Ponce-Alvarez et al., 2015). A critical next step in our understanding is to evaluate not only the degree of metastability arising spontaneously from the brain's intrinsic network dynamics but also the degree of metastability engendered by the attendant demands of a task (Fingelkurts & Fingelkurts, 2001; Rabinovich et al., 2008). Here, we invoke the theoretical framework of metastable coordination dynamics (Kelso, 1995, 2012) to explain how resting state networks are dynamically linked into task-dependent neurocognitive networks (Bressler & Kelso, 2016). Given that patterns of brain activity appear to be more stable during cognitive operations requiring explicit attention (Chen et al., 2015; Cohen, 2018; Elton & Gao, 2015; Hutchison & Morton, 2015), we anticipated reduced metastability between task-relevant neural networks as a function of task performance.
In light of the foregoing, we tested the hypothesis that coupling between the brain's large-scale networks is more metastable at rest than during the execution of an explicit task. We compared the metastability of fMRI BOLD signal in resting and task-evoked functional MRI data in a large cohort of healthy individuals (N = 566) from the Human Connectome Project (Van Essen et al., 2013). Changes in metastability were sought among 13 resting state networks encompassing hundreds of regions and every major brain system (Gordon et al., 2016). Finally, a link between the metastability of individual network connections and task performance was sought across several cognitive domains.
The methodological analysis comprised five stages: (1) changes in metastability between large scale networks evoked by seven types of task-based reasoning were examined using the network based statistic (NBS); (2) seven tasks (plus rest) were classified according to the metastability exhibited between large-scale brain networks using a convolutional neural network (CNN); (3) changes in metastability common to all seven tasks were sought through principal component analysis (PCA); (4) the cognitive relevance of metastability between networks was examined at rest and during the performance of an explicit task and (5) the contribution of metastable network dynamics to the efficient transformation between rest and task-driven network architectures, as captured by the similarity of task and rest-based network configurations, was investigated.
Overall, we found that–contrary to expectations–the metastability of couplings between large-scale networks was actively enhanced by task performance, principally in regions known to be devoted to cognitive control. Although metastability was evoked by task, cognitive performance was more closely aligned with the metastability of the brain's intrinsic network dynamics.
2 METHODS
2.1 Participants
Data were obtained through the Washington University-Minnesota Consortium Human Connectome Project (HCP; Van Essen et al., 2013). Subjects were recruited from Washington University and surrounding area. The present paper used 566 subjects from the 1,200 healthy young adult release (aged 22–35; see https://www.humanconnectome.org/data). All participants were screened for a history of neurological and psychiatric conditions and use of psychotropic drugs, as well as for physical conditions or bodily implants. Diagnosis with a mental health disorder and structural abnormalities (as revealed by fMRI) were also exclusion criteria. All participants supplied informed consent in accordance with the HCP research ethics board. The subset of subjects comprising monozygotic and dizygotic twin pairs were excluded from the present study. All subjects attained at a minimum a high school degree.
2.2 MRI parameters
In all parts of the HCP, participants were scanned on the same equipment using the same protocol (Smith et al., 2013). Whole-brain echoplanar scans were acquired with a 32 channel head coil on a modified 3T Siemens Skyra with TR = 720 ms, TE = 33.1 ms, flip angle = 52°, BW = 2,290 Hz/Px, in-plane FOV = 208 × 180 mm, 72 slices, 2.0 mm isotropic voxels, with a multi-band acceleration factor of 8. Rest (eyes open with fixation) and task-based fMRI data were collected over two sessions. Each session consisted of two rest imaging sessions of approximately 15 min each, followed by task-based acquisitions of varying length (totalling 30 min). The sessions were conducted on consecutive days at approximately the same time. Four tasks were acquired in the first session and three in the other. Except for the run duration, task-based data were acquired using the same EPI pulse sequence parameters as rest. Seven tasks lasting a total of 1 hr were acquired. Counter balancing of task order was not performed. The seven tasks (run times in minutes) were collected in the following order: working memory (10:02), gambling/reward learning (6:24), motor responses (7:08), language processing (7:54), social cognition (theory of mind; 6:54), relational reasoning (5:52) and emotion perception (4:32; Barch et al., 2013). Only resting state data from the first session were utilised. High-resolution 3D T1-weighted structural images were also acquired with the following parameters: TR = 2,400 ms, TE = 2.14 ms, TI = 1,000 ms, flip angle = 8°, BW = 210 Hz/Px, FOV = 224 × 224 and 0.7 mm isotropic voxels.
2.3 Task protocols
- Emotion perception: participants were presented with blocks of trials asking them to decide which of two faces presented on the bottom of the screen matched the face at the top of the screen, or which of two shapes presented at the bottom of the screen matched the shape at the top of the screen. The faces had either an angry or fearful expression (Hariri, Tessitore, Mattay, Fera, & Weinberger, 2002).
- Relational reasoning: participants were presented with two pairs of objects, with one pair at the top of the screen and the other pair at the bottom of the screen. Subjects were first asked to decide if the top pair of objects differed in shape or differed in texture and then to decide whether the bottom pair of objects also differed along the same dimension (Smith, Keramatian, & Christoff, 2007).
- Language processing: the task comprised a story and math component. The story blocks presented participants with brief auditory stories (5–9 sentences) adapted from Aesop's fables, followed by a 2-alternative forced-choice question that asked participants about the topic of the story. The math task also presented trials auditorily and required subjects to complete addition and subtraction problems (Binder et al., 2011).
- Two-back working memory: task participants were presented with blocks of trials that consisted of pictures of places, tools, faces and body parts (non-mutilated, non-nude). The task consisted of indicating when the current stimulus matched the one from two steps earlier.
- Gambling/reward learning: participants were asked to guess the number on a mystery card in order to win or lose money. Participants were told that potential card numbers ranged from one to nine and that the mystery card number was more than or less than five (Delgado, Nystrom, Fissell, Noll, & Fiez, 2000).
- Social cognition (theory of mind): participants were presented with short video clips (20 s) of objects (squares, circles, triangles) that either interacted in some way, or moved randomly on the screen. After each video clip, participants were asked to judge whether a mental interaction had occurred; did the shapes appear to take into account each other's thoughts and feelings? (Castelli, Happé, Frith, & Frith, 2013; Wheatley, Milleville, & Martin, 2007).
- Motor responses: participants were presented with visual cues that asked them to either tap their left or right fingers, squeeze their left or right toes, or move their tongue (Buckner, Krienen, Castellanos, Diaz, & Yeo, 2011; Yeo et al., 2011).
Full timing and trial structure for the seven tasks are provided as Data S1.
2.4 Task fMRI behavioural data
Task performance was evaluated using behavioural accuracy and reaction time data. Only those tasks which showed normally distributed behavioural accuracy scores were subject to linear regression analyses (see Figure S1). As confirmed by a Kolmogorov–Smirnov test (p < .05) three of the seven tasks satisfied this criterion including relational reasoning (M = 0.76, SD = 0.12), language processing (M = 0.88, SD = 0.71) and working memory (M = 0.83, SD = 0.10). Of the other four tasks, the gambling task accuracies were no better than chance (participants were asked to guess if a mystery card was higher or lower than five). The emotion (M = 0.97, SD = 0.03) and social (M = 0.96, SD = 0.12) task accuracies were perfect or near perfect for most subjects and hence showed a strong ceiling effect. Finally, the motor task accuracy scores were not recorded (subjects were asked to move tongue, hands, or feet). All seven tasks showed normally distributed reaction time data, as confirmed by a one-sample Kolmogorov–Smirnov test (p < .05).
2.5 Cognitive measures
- Penn progressive matrices (PMAT): measures fluid intelligence via non-verbal reasoning using an abbreviated version of the Raven's Progressive Matrices Form A developed by Gur and colleagues (Bilker et al., 2012). Participants are presented with patterns made up of 2 × 2, 3 × 3 or 1 × 5 arrangements of squares, with one of the squares missing. The participant must pick one of five response choices that best fits the missing square on the pattern. The task has 24 items and three bonus items, arranged in order of increasing difficulty. However, the task discontinues if the participant makes five incorrect responses in a row.
- NIH Toolbox Picture Vocabulary Test and NIH Toolbox Oral Reading Recognition Test: measures crystallised intelligence by averaging the normalised scores of each of the Toolbox tests that are crystallised measures, then derives scale scores based on this new distribution. One can interpret this crystallised cognition composite as a more global assessment of individual and group verbal reasoning. Higher scores indicate higher levels of functioning. The Picture Vocabulary Test is a measure of general vocabulary knowledge for ages 3–85. The participant is presented with an audio recording of a word and four photographic images on the computer screen and is asked to select the picture that most closely matches the meaning of the word. Higher scores indicate higher vocabulary ability. The Reading Test is a measure of reading ability for ages 7–85. The participant is asked to read and pronounce letters and words as accurately as possible. Higher scores indicate better reading ability.
- NIH Toolbox Flanker Inhibitory Control and Attention Test: measures executive function, specifically tapping inhibitory control and attention for ages 3–85. The test requires the participant to focus on a given stimulus whilst inhibiting attention to flanking stimuli. Sometimes the middle stimulus points in the same direction as the flankers (congruent) and sometimes in the opposite direction (incongruent). Scoring is based on a combination of accuracy and reaction time.
All three cognitive measures were consistent with a normal distribution as confirmed by a one-sample Kolmogorov–Smirnov test (p < .05).
2.6 fMRI pre-processing
All pre-processing was conducted using custom scripts developed in MATLAB 2017a (The MathWorks, Inc., Natick, MA). Motion between successive frames was estimated using framewise displacement (FD) and root mean square change in BOLD signal (DVARS; Power, Barnes, Snyder, Schlaggar, & Petersen, 2012; Power et al., 2014; Power, Schlaggar, & Petersen, 2015; Burgess et al., 2016). FD was calculated from the derivatives of the six rigid-body realignment parameters estimated during standard volume realignment. In keeping with previous time-resolved experiments (Shine, 2016a), and in light of the significant positive relationship between motion during rest and motion during each of the seven tasks (p < .01), if more than 20% of a subject's resting state frames exceeded FD > 0.5 mm they were excluded from further analysis. Based on these criteria, 566 out of 890 subjects were retained for further analysis. To preserve the temporal structure of the signal no data 'scrubbing' was performed (Power, 2014). Motion was defined as the SD in FD. A positive relationship between motion during rest and motion during task was identified in each of the seven domains. These included emotion perception (F[1,565] = 21.6, p < .01, r = .34), relational reasoning (F[1,565] = 43.6, p < .01, r = .28), language processing (F[1,565] = 48.2, p < .01, r = .37), working memory (F[1,565] = 107.1, p < .01, r = .42), gambling/reward learning (F[1,565] = 57.2, p < .01, r = .35), social cognition (F[1,565] = 96.8, p < .01, r = .46) and motor responses (F[1,565] = 62.9, p < .01, r = .31).
We used a minimally pre-processed version of the data that included spatial normalisation to a standard template, motion correction, slice timing correction, intensity normalisation and surface and parcel constrained smoothing of 2 mm full width at half maximum (Glasser et al., 2013). The data corresponded to the standard 'grayordinate' space consisting of left and right cortical surface meshes and a set of subcortical volume parcels which have greater spatial correspondence across subjects than volumetrically aligned data (Glasser et al., 2016). To facilitate comparison between rest and task-based conditions both sets of data were identically processed.
Cortical reconstruction and volumetric segmentation was performed with the Freesurfer image analysis suite, which is documented and freely available for download online (http://surfer.nmr.mgh.harvard.edu/). Briefly, this processing includes motion correction and averaging (Reuter, Rosas, & Fischl, 2010) of multiple volumetric T1 weighted images (when more than one is available), removal of non-brain tissue using a hybrid watershed/surface deformation procedure (Ségonne et al., 2004), automated Talairach transformation, segmentation of the subcortical white matter and deep grey matter volumetric structures (including hippocampus, amygdala, caudate, putamen, ventricles; Fischl et al., 2002, 2004) intensity normalisation (Sled, Zijdenbos, & Evans, 1998), tessellation of the grey matter white matter boundary, automated topology correction (Fischl, Liu, & Dale, 2001; Ségonne, Pacheco, & Fischl, 2007) and surface deformation following intensity gradients to optimally place the grey/white and grey/cerebrospinal fluid borders at the location where the greatest shift in intensity defines the transition to the other tissue class (Dale, Fischl, & Sereno, 1999; Dale & Sereno, 1993; Fischl and Dale, 2000). To mitigate partial volume effects, white matter and ventricle masks were subsequently eroded by one voxel on all edges using FSL (FMRIB's Software Library, www.fmrib.ox.ac.uk/fsl) tool fslmaths (Jenkinson, Beckmann, Behrens, Woolrich, & Smith, 2012; Smith et al., 2004; Woolrich et al., 2009). Average signals were extracted from the voxels corresponding to the ventricles and white matter anatomy. Variables of no interest were removed from the time series by linear regression. These included six linear head motion parameters, mean ventricle and white matter signals, and corresponding derivatives.
To obtain meaningful signal phases and avoid introducing artefactual correlations, the empirical BOLD signal was bandpass filtered (Glerean et al., 2012). Since low frequency components of the fMRI signal (0–0.15 Hz) are attributable to task-related activity whereas functional associations between high frequency components (0.2–0.4 Hz) are not (Sun, Miller, & D'Esposito, 2004), a temporal bandpass filter (0.06–0.125 Hz) was applied to the data (Shine et al., 2016a). The frequency range 0.06–0.125 Hz is thought to be especially sensitive to dynamic changes in task-related functional brain architecture (Bassett et al., 2011, 2013; Bassett, Yang, Wymbs, & Grafton, 2015; Glerean et al., 2012).
Since each task comprised two runs (one from each session) both runs were concatenated into a single time series. The individual signals were demeaned and normalised by z-scoring the data. To pre-empt the possibility that variation in synchrony (our definition of metastability) was being driven by alternating blocks of task and fixation, task blocks were concatenated. Since artificially concatenating a series of disjoint task blocks resulted in a discontinuous time series, the analysis was also performed with cue and fixation blocks included. Overall, retaining cue and fixation blocks did not alter the pattern of metastability between large-scale networks (only the statistical significance). The present analysis pertains to the case where cue and fixation blocks are removed. To ensure that any observed differences were due to dynamics rather than bias associated with signal length, the same number of contiguous frames from task and rest were utilised; the resting state scan was truncated to match the length of the task run (after cue and fixation blocks were removed).
2.7 Brain parcellation
Mean time series were extracted from regions of interest defined by the Gordon atlas (Gordon et al., 2016). The separation of regions into functionally discrete time courses is especially suitable for interrogating dynamic fluctuations in synchrony between large-scale networks. The Gordon atlas assigns regions to one of 12 large-scale networks corresponding to abrupt transitions in resting state functional connectivity. These include dorsal attention, ventral attention, fronto-parietal, cingulo-opercular, salience, default mode, medial parietal, parietal-occipital, visual, motor mouth, motor hand and auditory networks. Regions outside these domains are labelled as 'none'. The atlas was downloaded from the Brain Analysis Library of Spatial Maps and Atlases database (https://balsa.wustl.edu; Van Essen et al., 2017). Whole-brain coverage consisted of 333 cortical regions (161 and 162 regions from left and right hemispheres respectively), and one subcortical volume corresponding to the thalamus. The thalamus, which exhibits domain-general engagement across multiple cognitive functions, also plays a critical role in integrating information across functional brain networks (Hwang, Bertolero, Liu, & D'Esposito, 2017).
2.8 Calculating resting state network metastability

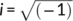 . If Bedrosian's theorem (Bedrosian, 2008) is respected then the analytic signal of a time series can be rewritten as,
. If Bedrosian's theorem (Bedrosian, 2008) is respected then the analytic signal of a time series can be rewritten as,

To relate findings to the existing literature and to preserve potentially important task-based neural components in the data, global signal regression was not performed in this study. The global signal is defined as the mean time-series of signal intensity across all voxels. Its removal by inclusion as a nuisance regressor in the general linear model, or global signal regression, has provoked controversy (Liu, Nalci, & Falahpour, 2017; Murphy & Fox, 2017). On the one hand, the global signal represents a 'catch-all' component reflecting various respiratory, scanner and motion related artefacts (Liu et al., 2017; Murphy & Fox, 2017). On the other hand, its removal has been linked to the introduction of artefactual anti-correlations in the data (Fox, Zhang, Snyder, & Raichle, 2009; Murphy, Birn, Handwerker, Jones, & Bandettini, 2009), altered distributions in regional signal correlations (Gotts et al., 2013) and distorted case–control comparisons of functional connectivity measures (Gotts et al., 2013). Moreover, although the global signal reflects non-neural confounds in the data, it may also contain a substantial neural component. This is supported by evidence suggesting that spontaneous fluctuations in local field potentials correlate with fMRI signals across the entire cerebral cortex (Schölvinck, Maier, Ye, Duyn, & Leopold, 2010) and by a recent study suggesting that 14–50% of the variance of the global signal is related to network-specific time series and not to factors such as arousal (Gotts, Gilmore, & Martin, 2020). These findings, combined with studies demonstrating a behavioural connection (Chang et al., 2016; Wen & Liu, 2016; Wong, Olafsson, Tal, & Liu, 2013), suggest that global signal regression is removing important neural information. Ultimately, whether or not to perform global signal regression is contingent upon the scientific aims of the investigation and must be taken into account when interpreting the results (Liu et al., 2017; Murphy et al., 2009). The effect of removing the global signal on estimates of metastability is provided as a supplemental analysis (see Figure S2).
The Hilbert transform is able to produce an analytic signal with meaningful envelope and phase for any finite block of bandpass filtered data. Issues may arise when attempting to extract meaningful phase relationships from short time series however, as robust estimation of metastability is contingent on time series being of sufficient length so as to permit adequate variation in synchrony between a set of regions to occur. The impact of estimating metastability on time series of different lengths is provided as supplementary information (see Figure S3). Estimates of metastability were stable across the length of time series considered in the present study including the shortest, emotion (4:32) and the longest, rest (15 mins). Sampling rate was not considered a significant factor in the estimation of metastability due to the already slow haemodynamic response.

The Hilbert transform permits temporal relationships between brain regions to be analysed by extracting the phase of a signal and discarding its amplitude. Accordingly, the result of the Hilbert transform may be conceptualised as a set of phase oscillators rotating around the circumference of the unit circle. The length of the vector pointing from the centre of the circle to the mean phase of the group is commensurate with the synchrony between oscillators. Variation in the length of this vector over time, from zero (no synchrony) to one (full synchrony), provides a measure of the system's overall metastability. Although two systems may differ in terms of synchrony, as reflected by vectors of different lengths, both can share the same metastability as reflected by a vector of constant length over time. The situation is illustrated by two oscillators rotating in-phase (with synchrony equal to one) versus two oscillators rotating  out-of-phase (with synchrony equal to 0.5). In both cases, the lengths of the vectors is constant and metastability is zero.
out-of-phase (with synchrony equal to 0.5). In both cases, the lengths of the vectors is constant and metastability is zero.
2.9 Assessing changes in empirical resting state network metastability during tasks
Given that (1) the current formulation of metastability permits calculation between a group of regions; and (2) interconnected subnetworks convey more behaviourally relevant information than functional connections observed between pairs of regions observed in isolation, we advocate for a method that exploits the clustering structure of connectivity alterations between functionally related networks. For this reason, we applied the NBS to estimates of empirical metastability obtained from fMRI data at the network rather than regional level (see also, Alderson et al., 2018). For each subject, we estimated an 'interaction matrix' reflecting the metastable interactions of the 13 resting state networks (and thalamus) defined by the Gordon atlas. The same procedure was applied to compute an equivalent interaction matrix based on synchrony.
The NBS is a non-parametric statistical test designed to deal with the multiple comparisons problem by identifying the largest connected sub-component (either increases or decreases) in topological space whilst controlling the family wise error rate (FWER). To date, several studies have used the method to identify pairwise regional connections that are associated with either an experimental effect or between-group difference in functional connectivity (Zalesky, Fornito, & Bullmore, 2010). Here, we use the NBS to identify topological clusters of altered metastability (or synchrony) between empirical resting state networks under different conditions of task relative to rest.
Mass univariate testing was performed at every connection in the graph to provide a single test statistic supporting evidence in favour of the null hypothesis, namely, no statistically significant difference in the means of resting state and task-based metastability. The test statistic was subsequently thresholded at an arbitrary value with the set of supra-threshold connections forming a candidate set of connections for which the null hypothesis was tested. Topological clusters were identified between the set of supra-threshold connections for which a single connected path existed. The null hypothesis, therefore, was accepted or rejected at the level of the entire connected graph rather than at the level of an individual network connection. The above steps were repeated in order to construct an empirical null distribution of the largest connected component sizes. Finally, FWE-corrected p values, corresponding to the proportion of permutations for which the largest component was of the same size or greater, were computed for each component using permutation testing.
2.10 Classification of task and rest data
The interaction matrices corresponding to the seven different tasks (plus rest) were classified using a modified CNN architecture (Figure 1). BrainNetCNN is the first deep learning framework designed specifically to leverage the topological relationships between nodes in brain network data, outperforming a fully connected neural network with the same number of parameters (Kawahara et al., 2017). The architecture of BrainNetCNN is motivated by the understanding that local neighbourhoods in connectome data are different from those found in traditional datasets informed by images. Patterns are not shift-invariant (as is a face in a photograph) and the features captured by the local neighbourhood (e.g., a 3 × 3 convolutional filter) are not necessarily interpretable when the ordering of nodes is arbitrary.

To reduce the number of parameters we included only a single edge-to-edge layer (Meszlényi, Buza, & Vidnyánszky, 2017). The input to the CNN is the set of 14 × 14 interaction matrices that capture the metastability/synchrony between the 13 resting state networks (plus the thalamus) defined by the Gordon atlas. The network classifies the data into one of the seven tasks or the subject's resting state (random classification accuracy is 12.5%). The model was evaluated using k-fold cross validation where k = 5. This value of k has been shown to yield test error rate estimates that suffer neither from excessively high bias nor from very high variance (Kuhn & Johnson, 2013). The original dataset was partitioned randomly into training (60%), validation (20%) and testing sets (20%). That is, 340 subjects were assigned for training the model, 113 subjects were assigned for tuning the model's hyperparameters, and a further 113 were withheld for validating the performance of the trained model. This was repeated five times for k-fold using different test subsets each time. In the case of metastability, each of the 566 subjects was associated with eight interaction matrices (seven task-based interaction matrices and one resting state interaction matrix). The same was true in the case of synchrony. Performance was evaluated using classification accuracy, that is, the proportion of correctly identified instances. The above procedure was repeated twice, once for the interaction matrices capturing metastability and again using the interaction matrices based on synchrony.
The CNN was implemented in Python using the Pytorch framework (Paszke et al., 2017). Rectified linear units (RELUs; Nair & Hinton, 2010) were used as activation functions between layers and the probability of each class was calculated at the output layer using the soft max function (Bridle, 1990). The network was trained using the Adam optimiser (Kingma & Ba, 2015) with mini-batch size of 128, a learning rate of 0.001 and momentum of 0.9. Dropout regularisation of 0.6 was applied between layers to prevent over-fitting (Srivastava, Hinton, Krizhevsky, Sutskever, & Salakhutdinov, 2014; Wager, Wang, & Liang, 2013). The model minimised a cost function associated with the cross-entropy loss. Hyperparameters used in the optimisation stage included momentum and drop-out regularisation.
2.11 Defining update/reconfiguration efficiency
The ability to switch from a resting state network architecture into a task-based configuration was designated as update efficiency (Schultz & Cole, 2016). Highly similar rest and task-based network configurations are commensurate with high update efficiency, as few changes are required to transition between the two whilst highly dissimilar resting state and task-based architectures are linked to low update efficiency, reflecting the many changes that are required to make the switch. Update efficiencies were calculated for all 566 subjects by vectorising the upper triangular half and diagonal of the rest and task-general interaction matrices and calculating their Pearson's correlation coefficients. The latter were converted to a normal distribution by performing Fisher's z transformation.
3 RESULTS
3.1 Higher global metastability during task than rest
The metastability of fMRI BOLD signal was examined during the resting state and during the execution of several cognitively demanding tasks (Figure 2). One-way ANOVA identified a statistically significant difference between groups (F[7,4,520] = 37.32, p < .01). Subsequent Tukey post hoc test revealed significantly higher global metastability during the seven tasks as compared to the resting state (M = 0.112, SD = 0.019). These included emotion perception (M = 0.132, SD = 0.031), relational reasoning (M = 0.133, SD = 0.030), language processing (M = 0.134, SD = 0.027), working memory (M = 0.135, SD = 0.027), gambling/reward learning (M = 0.136, SD = 0.029), social cognition (M = 0.141, SD = 0.029) and motor responses (M = 0.143, SD = 0.030). Changes in metastability are compared among the seven tasks as a supplemental analysis.

3.2 Task related increases in metastability between resting state networks
The NBS was subsequently used to identify changes in the metastability of fMRI BOLD signal of individual network connections (task vs. rest). In total, 566 resting state interaction matrices were compared to 566 task-based interaction matrices within each of the seven behavioural task domains. The null hypothesis, namely, that there was no difference in metastability between rest and task, could then be rejected at the level of individual network connections.
Consistent with the role of resting state networks in mediating behaviour (Sadaghiani, 2010; Sadaghiani & Kleinschmidt, 2013), the NBS identified statistically significant (p < .01; corrected) increases in metastability between several large-scale networks during task engagement relative to the more unconstrained resting state. A single test statistic threshold, that is, 16, was selected for all seven tasks so as to permit visualisation of the increases in metastability between large-scale networks on the same scale (Figure 3). Figure 3 shows the largest connected sub-graph of increased metastability detected by the NBS at a fixed threshold for all seven tasks where each node is scaled to reflect its relative importance within the sub-graph (the sum of its effect sizes).

To characterise these increases across all seven tasks, we summed the test statistics associated with each network's interactions (see Figure S4). Networks related to cognitive control, including the fronto-parietal, dorsal attention and cingulo-opercular networks, along with thalamo-cortical networks linked to memory, learning and flexible adaptation (Alcaraz et al., 2018; Wolff & Vann, 2018), demonstrated the most consistent increases in metastability across the seven tasks. Moreover, metastability increased to a greater extent in regions associated with cognitive control than regions linked to sensory–motor processing (see Figures S5–S9).
3.3 Increases in metastability were more widespread than equivalent increases in synchrony
Edges associated with each sub-graph (metastability and synchrony) were summed to reveal the total number of network connections recruited for each task (Figure 4). Increases in metastability spanned a greater number of cognitive subsystems than equivalent increases in synchrony (in all but emotion and motor tasks). In both cases, the NBS received an identical threshold.

3.4 Each task is characterised by a small number of task-evoked changes in synchrony
The highly correlated properties of metastability and synchrony (see Figure S10) were disassociated using a deep learning framework. Accordingly, task and rest network states captured by the 14 × 14 interaction matrices of metastability and synchrony were provided as input to a CNN for classification. The network correctly identified the seven different tasks (plus rest) based on synchrony with high accuracy (76% average; chance level 12.5%; Figure 5a) but performed less well when trained on metastability (46% average; Figure 5b). The high sensitivity (true positive rate) and specificity (true negative rate) exhibited by the classifier when trained on synchronous interactions between networks suggests that each behavioural domain was characterised by a small number of unique task-evoked network changes. This was confirmed by masking out inputs (interactions between networks) relevant for correct classification (Figure 6) and re-evaluating the pre-trained classifier. Overall, occluded inputs were associated with exceptionally poor classification accuracy (Figure 5c).


In detail, guided backpropagation was used to identify the most important inputs for correctly classifying each task. Guided backpropagation provides a set of gradients relating input to output. High gradients at the input level have a large effect on the output and are therefore important for classification. Recall, that each row/column of the 14 × 14 input represented the interaction of a single network (plus thalamus) with 14 others. Thus, guided backpropagation produced a 14 × 14 matrix of gradients. A consensus across all subjects for a particular task was obtained by setting each subject's top 10% of gradients (the most positive gradients) to one and the remaining entries to zero, summing the matrices and dividing by the total number of subjects. Entries important for correct classification in more than 90% of individuals were set to zero in the input (the interaction matrix). The performance of the pre-trained classifier was then re-evaluated based on the occluded input data. In a separate experiment, retraining on the occluded inputs also produced extremely poor classification accuracy (not shown).
3.5 Different behaviours recruit a similar set of metastable connections
So far, we have demonstrated that increases in metastability can be distinguished from increases in synchrony in two ways: (1) their overall network size, that is, increases in metastability encompass a wider network of cognitive-related brain systems than those based on synchrony; and (2) their discriminatory utility, that is, tasks can be identified with high accuracy based on a small subset of network changes in synchrony (but much less so in terms of metastability). Taken as a whole, these findings led us to hypothesise that commonalities between tasks may be centred on a metastable core of task-general network interactions.
To quantify the degree to which the seven task-based configurations shared features in common, we used a dimension-reduction tool–PCA–to reduce a larger set of variables (the seven task-based interaction matrices) to a smaller set (a single task-general network architecture) retaining most of the information. Accordingly, entering the seven task-based interaction matrices based on metastability into a PCA yielded a single task-general architecture for each subject. On average, the first principal component accounted for 78% of the variance between the seven tasks. The loadings were positive and distributed equally between the seven tasks suggesting that each task-based configuration contributed equally to the task-general structure. These included emotion perception = 0.37, relational reasoning = 0.37, language processing = 0.36, working memory = 0.36, gambling/reward learning = 0.36, social cognition = 0.37 and motor responses = 0.38. This result likely speaks to the high similarity between task-based configurations engaged by different behaviours (see Figure S11). An exemplar task-general architecture was subsequently derived through simple averaging across subjects.
3.6 Task general architecture is composed of subnetworks of increased and decreased metastability
We explicated this structure by performing another PCA analysis on the interaction matrices obtained by subtracting task from rest. In doing so, the task general architecture was decomposed into subnetworks of high (Figure 7; top) and low (Figure 7; middle) metastability. High metastability (Figure 7; red) was found in networks associated with cognitive control including dorsal attention (selective attention in external visuospatial domains) and fronto-parietal networks (adaptive task control). Tertiary thalamo-cortical contributions were also apparent (memory, category learning and adaptive flexibility). In contrast, low metastability (Figure 7; blue) was linked to unimodal (or modality specific) sensory processing architecture including motor, auditory and visual networks.

3.7 High metastability of cognitive control systems at rest is predictive of task performance
We next examined the metastable interactions of large-scale networks during task engagement for evidence that they informed behaviour. Behavioural accuracy scores for each subject were entered into a linear regression analysis as the dependent variable with one of 196 (14 × 14) task-based network connections (estimated in terms of metastability) as predictors. Overall, metastable interactions between networks during task did not explain the variance in cognitive ability. Resting state metastability was then entered as an additional independent factor. Across the three in-scanner tasks, several network connections demonstrated a significant positive association between intrinsic metastability and cognitive performance (Figure 8 bottom; p < .01; FDR corrected for multiple comparisons). Three additional cognitive measures acquired outside the scanner were also analysed. These included fluid intelligence, crystallised intelligence and executive function. Across the three measures, several network connections demonstrated a significant positive association between intrinsic metastability and cognitive performance (Figure 8 top; p < .01; FDR corrected for multiple comparisons). The addition of reaction time data across the six tasks did not increase the percentage of explained variance in cognitive performance. In a separate linear regression analysis, no statistically significant associations between behaviour/cognition and network synchrony were identified (p < .01; FDR corrected for multiple comparisons).

Interactions between networks are presented as circular graphs where each edge represents a significant positive correlation between network metastability and cognition (Figure 8). In the main, the metastability of large-scale networks related to cognitive control was strongly related to task performance (see also Figures S12 and S13). These included fronto-parietal, dorsal attention and cingulo-opercular networks, and to a lesser extent the default mode, ventral attention and salience networks. In general, the metastability of networks related to primary sensory processing (including motor, auditory and visual networks) was less relevant to cognitive performance (see again Figures S12 and S13). These results are broadly consistent with the task-general network architecture previously discussed.
Concerning the three tasks assessed outside the scanner, the metastability of connections associated with cognitive control networks, including fronto-parietal, cingulo-opercular and dorsal attention, was strongly related to fluid intelligence. In contrast, the metastability of these networks was less important in the execution of crystallised intelligence (see Figure S14 for a statistical comparison). A qualitative comparison of these same tasks revealed an important role of cingulo-opercular network metastability in executive function/inhibitory control. In regard to the three in-scanner tasks, the working memory task was qualitatively distinguished from relational reasoning and language processing by a stronger association between metastability and behavioural accuracy in the thalamus. Likewise, language processing was qualitatively distinguished from relational reasoning and working memory by virtue of a stronger association between metastability and cognition in the connectivity of the ventral attention network. All six behavioural domains, including relational reasoning, language processing, working memory, fluid intelligence, crystallised intelligence and executive function/inhibitory control, demonstrated significant positive correlations between cognitive performance and spontaneous metastability in regions associated with cognitive control (including the fronto-parietal, dorsal attention and cingulo-opercular networks).
3.8 High metastability within (and between) cognitive control systems at rest promotes efficient switching into task
Even though it is self-evident that for high update efficiency to be achieved, a subject's resting and task-general architectures must be in agreement, not all of these relationships will necessarily hold statistically at the chosen significance level (p < .01; FDR). For this reason, we correlated the metastability of individual network connections with update efficiency and obtained the slope of the regression equation (Figure 9a) and its significance (Figure 9b).

A significant positive relationship between update efficiency and metastability was found in the fronto-parietal, dorsal attention, cingulo-opercular and default mode networks (Figure 9c; right; red), and a significant negative relationship in the motor, auditory and visual networks (Figure 9c; left; blue; p < .01; FDR corrected for multiple comparisons). As expected, these results are in accord with the structure of the task-general configuration (Figure 7). Overall, high update efficiency was characterised by dynamic flexibility in the connectivity of networks implicated in cognitive control and dynamic stability in primary sensory networks. Some networks such as the salience, ventral attention, medial parietal, parieto-occipital and thalamus were sites of convergence for both high and low metastability connections (Figure 9c; centre; yellow nodes).
3.9 Subjects with similar resting and task-general architectures demonstrate superior performance
We next examined whether cognitive performance and the efficiency of the transformation between rest and task-based neural architectures are related (Schultz & Cole, 2016). To this end, behavioural accuracies and update efficiencies were entered into linear regression analysis. Statistically significant (p < .05; FDR corrected for multiple comparisons) relationships between performance and update efficiency were identified in the three in-scanner tasks. These included relational reasoning (F[1,564] = 5.0, p = .026, r = .21), language processing (F[1,564] = 5.3, p = .021, r = .24) and working memory (F[1,564] = 4.9, p = .027, r = .20). Since metastability and update efficiency are correlated (in some connections) and since update efficiency predicts performance, we also confirmed that the efficiency-performance relations survived in the face of controlling for global metastability. Overall, these findings suggest that cognitive ability is contingent upon resting state architecture being similar or 'pre-configured' to a task-general arrangement.
4 DISCUSSION
The present paper set out to answer a relatively simple question: is metastable neural dynamics higher at rest or during the performance of an explicit task? We sought to answer this question by comparing the metastability of the brain's large-scale networks at rest and during the execution of several cognitively demanding tasks. Current theory suggests that spontaneous neural dynamics represent a repository of functional states from which more stable global brain states are constructed during task-based reasoning. Metastability between networks should therefore be maximal when subjects are at 'cognitive rest' and diminished during times of heightened cognitive demand. Surprisingly, our findings support an alternative possibility: metastability (or dynamic flexibility) between neural networks is actively enhanced by task engagement (Figures 2 and 3). Explicit cognition was characterised by two types of network architecture: (1) a task-specific network structure based on a circumscribed set of task-evoked changes in synchrony (Figure 6) and (2) a task-general network structure based on widespread changes in metastability (Figure 7). This task-general architecture was principally characterised by increased metastability in cognitive control networks and decreased metastability in sensory regions (Figure 7). Although resting state networks were dynamically linked into context-dependent neurocognitive structures by virtue of task engagement, cognitive performance was more closely linked to the intrinsic activity of large-scale networks (Figure 8). High metastability in the intrinsic connectivity of cognitive control networks was associated with novel problem solving (so-called fluid intelligence) but was less important in tasks relying on previous experience (so-called crystallised intelligence). Critically, subjects with resting state architectures similar or 'pre-configured' to a task-general configuration demonstrated superior cognitive performance that was linked to efficient switching of network states (Figure 9). Overall, our findings suggest a key linkage between the intrinsic metastability of the brain's large-scale network connectivity and cognition.
The present study used one measure of connectivity–metastability–to assess the dynamic stability~flexibility within and between large-scale networks of the brain. Increased stability of dynamic functional connectivity appears to be a general property of cognitive engagement across multiple behavioural paradigms irrespective of the type of dynamic functional connectivity method employed (Cohen, 2018). Consistent with the properties of a critical system, dynamic functional connectivity is especially stable during tasks requiring sustained attention (Haimovici, Tagliazucchi, Balenzuela, & Chialvo, 2013; Meisel, Olbrich, Shriki, & Achermann, 2013). Focused cognition appears to induce the sub-critical dynamics necessary for reducing interference and optimising task performance (Fagerholm et al., 2015). Such dynamic stability is associated with increased integration across cognitive control networks and between cognitive control and other task-relevant networks (Chen, Chang, Greicius, & Glover, 2015; Elton & Gao, 2015; Hutchison & Morton, 2015). On the surface, increased stability between networks during task engagement appears at odds with the present finding of increased metastability. How can something be both flexible and stable at the same time? Fortunately, the paradox is resolved when we recognise that metastability and synchrony are correlated attributes of brain function. Thus, even though metastability (or the variation in synchrony) is increasing, so too is the average synchrony (which can be viewed as a measure of stability in this context). It should be emphasised that purely synchronous episodes of spatio-temporal coordination between networks is not reflective of typical neurocognitive processing. Our measure of synchrony reflects this reality by reporting the average synchrony (or mean phase coherence) over a period of time corresponding to a series of concatenated task blocks. Using this approach, we show that task blocks are characterised by on average higher levels of synchrony and higher levels of metastability simultaneously. That is, synchrony (the mean phase coherence over time) and metastability (the variation in the mean phase coherence over time) increase together–a behaviour observed in cortical slice cultures excited pharmacologically (Yang, Shew, Roy, & Plenz, 2012). In light of this, synchrony and metastability should be considered as related rather than contrary methods of viewing brain activity. Two possibilities exist: (1) that metastability is the price paid for higher on average synchrony, that is, it is a form of noise; or (2) metastability is an adaptive process reflecting the ongoing engagement and disengagement of neural systems relevant to cognitive and affective states. Our findings strongly support a functional view of metastability–cognitive performance is linked to the intrinsic metastability of large-scale networks. Indeed, theoretical and empirical findings suggest that intrinsic neural architecture is organised to support a range of functional states which can be a posteriori selected via exogenous input (Cabral, Kringelbach, & Deco, 2014; Deco et al., 2017; Deco & Kringelbach, 2016; Hansen, Battaglia, Spiegler, Deco, & Jirsa, 2014; Kringelbach et al., 2015; Ponce-Alvarez et al., 2015). One may argue that metastability is being driven by the variation in synchrony obtained by artificially concatenating a series of disjoint task blocks. However, in principle, metastability within a single task block should also be enhanced on the provision that synchrony also increases. Presently, the temporal limits imposed by fMRI and the relatively short task runs preclude a direct test of this hypothesis.
Task engagement was principally characterised by enhanced metastability in the connectivity of cognitive control networks (Figure 3). These included dorsal attention, fronto-parietal, cingulo-opercular, default mode and ventral attention networks. The present finding is consistent with the notion that task-positive networks correspond to areas of the brain that respond with activation increases to attention-demanding environments. The organisational state of the system may facilitate or inhibit the processing of incoming external stimuli by enacting a series of task-relevant synergies over the duration of the task (Kelso, 2009). Each of the networks presented similar but not identical patterns of engagement across the seven tasks. Two of the four, the dorsal attention and fronto-parietal networks, demonstrated consistent activity across all seven of the in-scanner tasks and were associated with the greatest increases in metastability. This raises the question of why dorsal fronto-parietal regions are involved in such a puzzling variety of tasks? One proposal is that cognitive computations relying on dorsal fronto-parietal areas are concerned with a single core function, namely 'offline motor planning' or 'action emulation' (Ptak, Schnider, & Fellrath, 2017). Such findings are consistent with the observation that hubs of the fronto-parietal network alter their pattern of functional connectivity with nodes of other networks based on goal-directed cognition in an adaptive domain-general manner (Cole et al., 2013; Cole, Repovš, & Anticevic, 2014). Rhythmic attentional sampling linked to theta-band activity in large-scale dorsal fronto-parietal regions (Fiebelkorn, Pinsk, & Kastner, 2018; Helfrich et al., 2018) may also account for increased metastability. Changes in metastability were not limited to interactions between cortical networks; they also involved subcortical, specifically, thalamo-cortical components. The finding of increased metastability between thalamus and dorsal fronto-parietal during task is consistent with empirical evidence–thalamic input wires the contextual associations upon which complex decisions depend into weakly connected cortical circuits (Halassa & Kastner, 2017; Schmitt et al., 2017).
The seven in-scanner tasks showed remarkably similar patterns of increased metastability between large-scale networks (Figure 3). Such findings resonate with prior studies showing similar patterns of static functional connectivity between different tasks (Cole, Bassett, et al., 2014; Schultz & Cole, 2016). Similarities between functional network configurations evoked under different behavioural paradigms constitute what has been referred to as a 'task-general architecture' (Cole, Bassett, et al., 2014; Schultz & Cole, 2016). Our findings suggest that different behaviours recruit a similar set of network connections through metastable neural dynamics. We base this claim on three observations: (1) increases in metastability were more widespread than increases in synchrony (Figure 4); (2) increases in synchrony were highly specific to each task, whereas increases in metastability were more task-general (Figures 5 and 6); and (3) most of the variance (78%) between tasks was accounted for by a single principal component (Figure 7). Our task-general configuration was organised into distinct regions of high and low metastability (Figure 7). Networks related to cognitive control exhibited dynamic flexibility whilst primary sensory networks favoured dynamic stability. These findings map onto our present understanding of cortical organisation and function. Primary sensory areas are responsible for processing a single modality whereas higher order association areas must integrate information into more complex representations, for example, for language, executive function, attention and memory. Thus, each step up the hierarchy entails integrating information from a greater diversity of sources, and this in turn is reflected in greater dynamic flexibility. From another perspective, widespread increases in metastability are quite surprising. They appear to indicate that most of the brain is involved during tasks which appears to contradict findings from cognitive fMRI studies showing only limited activation. One possibility is that some of these changes in metastability are linked to suppression of a given network during task performance.
Even during periods of apparent 'rest' a subject's network connectivity continued to display strong integrative and segregative tendencies linked to cognitive performance. Remarkably, a subject's ability to answer questions correctly, both in and out of the scanner, related to their intrinsic neural dynamics (Figure 8). Curiously, task-based changes in metastability were not related to cognitive performance, rather, subjects whose dynamics explored a greater range of network configurations at rest demonstrated the highest cognitive test scores and behavioural accuracy. Unlike crystallised intelligence, which was largely unrelated to metastability, the flexibility of network states afforded by metastable neural dynamics was strongly linked to fluid intelligence. Accumulating evidence suggests that human intelligence arises from the dynamic reorganisation of brain networks (Barbey, 2018). Flexible network transitions may support the 'difficult-to-reach' networks states associated with novel problem solving but may be less relevant for accessing the local knowledge and experience embodied in 'easy-to-reach' network configurations (Gu et al., 2015; Power & Petersen, 2013). The link between metastable neural dynamics and fluid intelligence was especially pronounced in systems involved in cognitive control. Our results are consistent with prior observations linking cognitive control capacity and fluid intelligence (Cole et al., 2012; Cole & Schneider, 2007; Conway, Cowan, Bunting, Therriault, & Minkoff, 2002).
Metastable network dynamics displayed a combination of trait- and state-like properties. Resting state metastability was significantly lower on Day 2 scanning sessions versus Day 1 scanning sessions (see Figure S15). This finding is consistent with metastability being linked to arousal or fatigue. Differences in metastability observed between scanning sessions (Day 1 vs. Day 2) was not sufficient to account for the observed increase in metastability across conditions (task vs. rest), as resting state metastability was significantly lower than task-driven metastability on both days. Interestingly, metastability at rest was proportional to the metastability exhibited during task in the same scanning session (see Figure S16) suggesting that metastability may also be linked to intrinsic factors such as anatomical connectivity; a view that is consistent with prior observations linking reduced metastability to altered network topology (Alderson et al., 2018; Córdova-Palomera et al., 2017; Hellyer et al., 2015; Váša et al., 2015).
So why is metastability at rest predictive of behavioural and cognitive performance as opposed to metastability during the task itself? The idea that functional couplings between regions at rest contain information relevant to cognition, perception and behaviour is supported by substantial empirical evidence (Sadaghiani, 2010; Sadaghiani & Kleinschmidt, 2013). Static resting state functional connectivity has been linked to a number of general cognitive abilities that include, among others, IQ, executive function, episodic memory and reading comprehension (for review, see Vaidya & Gordon, 2013). Thus, rather than simply reflecting invariant structural anatomy, historical co-activation patterns, or internal dynamics of local areas, intrinsic activity predicts subsequent perceptual processing (Busch, Dubois, & VanRullen, 2009; Hesselmann, Kell, Eger, & Kleinschmidt, 2008; Lou, Li, Philiastides, & Sajda, 2014; Mathewson, Gratton, Fabiani, Beck, & Ro, 2009; Sadaghiani, Hesselmann, & Kleinschmidt, 2009; Van Den Berg, Appelbaum, Clark, Lorist, & Woldorff, 2016; van Dijk, Schoffelen, Oostenveld, & Jensen, 2008).
Cognitive performance was also linked to the efficiency of the transformation between rest and task-based network architectures. Specifically, subjects whose resting state was similar to the task-general architecture garnered the highest cognitive scores. Successful cognition is likely contingent on possessing an adequate a priori dynamic configuration before the onset of task-relevant stimuli, as opposed to simple ad hoc adjustments after the fact (Bolt, Anderson, & Uddin, 2018). Thus, resting state activity may reflect the brain's predictive engagement with the environment (Clark, 2016; Sadaghiani, 2010; Sadaghiani & Kleinschmidt, 2013). Given that the resting state reflects previous experience and the anticipation of likely future events, a resting state network architecture 'pre-configured' to task is more in line with future cognitive requirements (Bar, 2011). Update efficiency, or the ability to switch from a rest- to a task-based configuration mapped onto our task-general architecture (Figure 9). High update efficiency was associated with dynamic flexibility in dorsal attention and fronto-parietal control networks and dynamic stability in primary sensory networks. Such findings conform to our intuitive expectation that cortical networks require varying amounts of dynamic flexibility to fulfil their function. Presumably, sensory networks continue to function even when subjects are cognitively at rest, hence, metastability increases less in these regions during times of heightened cognitive demand. Interestingly, several regions demonstrated high and low metastability connections. These included the salience network of which the insula–a known site of multi-modal integration of sensory, motor, emotional and cognitive information–is a part (Gogolla, 2017). Update efficiency based on a static measure of functional connectivity has been considered in a previous study (Schultz & Cole, 2016). Our work differs from this approach in that we consider the update efficiency within the context of a dynamic measure of network connectivity in which both integrative and segregative tendencies coexist (Kelso, 1995, 2012; Tognoli & Kelso, 2014b).
Potential limitations of the findings should be noted. Firstly, it is worth emphasising that the division of neural activity into intrinsic and task-evoked activity may be an artificial distinction and not an actual division respected by neural properties (Bolt et al., 2018). Moreover, the finding of increased metastability during task relative to rest may, to some extent, be dependent on factors related to experimental design including: (1) the time frame considered and (2) the temporal resolution of the imaging modality used. Increased metastability was identified in a series of concatenated task blocks. A more desirable experimental setup would compensate for fMRI's lack of temporal precision by measuring metastability over an extended period within a single task block. Doing so would ameliorate the potential confound of introducing variations in synchrony–our definition of metastability–by including rest, cue and fixation blocks. In the present paper, we mitigated this risk by estimating metastability with non-task block data removed. Moreover, due to restrictions on the length of the runs, metastability was estimated using the active and control components of the task. Hence, participants were not exclusively performing the 'cognitive task' but the 'control task' designed to compensate for non-specific effects (e.g., the zero back condition in the working memory task). It is worth noting however, that in most cases, there was either no control condition (e.g., the motor task) or subjects still performed a meaningful exercise in the control task condition (e.g., the language task which comprised a story and math component). Our results were robust in that metastability was not driven by bias associated with using time-series of different lengths (we used the same number of data points for rest and task), nor by alternating periods of rest and task within a single run (we removed cue and fixation blocks). Difference in acquisition moments (Day 1 vs. Day 2) also fail to account for the observed increase in metastability (rest vs. task) as resting state metastability was significantly lower than task-based metastability on both days. Despite attention to such factors, it remains of critical importance to establish the validity of observed synchronisation dynamics particularly in light of recent findings questioning the link between time varying functional connectivity and task-relevant neural information (Hindriks et al., 2016; Laumann et al., 2017; Liégeois, Laumann, Snyder, Zhou, & Yeo, 2017). Rapid changes in synchronisation may not be directly tied to external task demands, but rather to internally driven factors that include attention, motivation, arousal, fatigue, goals or levels of consciousness (Kucyi, 2017; Kucyi, Hove, Esterman, Hutchison, & Valera, 2017). For instance, network integration during rest is linked with greater pupil diameter; a proxy for arousal, as well as better task performance (Shine, et al 2016a, Bissett, et al., 2016). Similarly, across repeated resting state scans of the same individual, higher levels of fatigue are related to more stable estimates of dynamic functional connectivity whereas higher levels of attention are related to more variable measures of dynamic functional connectivity (Shine,et al., 2016b, Koyejo, & Poldrack, 2016). Rapid dynamics likely include both meaningful neural information and physiological signals that relate to differences in rate or volume of blood flow, respiration and heart rate (Cohen, 2018). In addition, movement of the subject within the scanner has been found to introduce spurious distance dependent correlations in resting state data (Power et al., 2012, 2014, 2015). Such motion induced artefacts arise through the introduction of spurious variance in the 'true' time series which decays with distance, thus, artefactual correlation is most similar in nearby voxels (Power et al., 2014). In a similar way, estimates of metastability may be biased by the introduction of spurious variations in synchrony between nearby regions. Thus, controlling for head motion related artifacts is also of central concern in the analysis of time-series phase estimates derived through the Hilbert transform. More aggressive motion control strategies beyond the traditional minimal pre-processing pipeline (regression of six head motion parameters, mean white matter and cerebrospinal fluid signal) may therefore be indicated (Ciric et al., 2017). Untangling the relationship between rapid dynamics, internal mentation, cognitive performance, affective states, physiological noise, arousal, fatigue and general cognition will become increasingly important as we seek to understand the neural basis of human behaviour. Finally, since task order was not counter balanced, the possibility that order effects may have influenced the task-based analyses cannot be discounted.
Taken together, our findings suggest that the metastable regime of coordination dynamics (Bressler & Kelso, 2016; Kelso, 1995, 2012; Tognoli & Kelso, 2014b) offers considerable potential as a theoretical and conceptual framework for linking resting state network activity to cognition and behaviour. Although resting state networks are dynamically reconfigured during cognitive engagement, task onset impacts the spatio-temporal properties of a pre-existing functional architecture. Such coordinative behaviour, organised through spontaneous metastable dynamics, appears to contribute to cognition by actively anticipating incoming stimuli. Cognitive function is therefore not primarily stimulus-driven or reflexive but arises from the brain's intrinsically self-organising character.
ACKNOWLEDGEMENTS
This work was supported by a Department for Economy, Northern Ireland PhD studentship. This work was also supported, in part, by a grant given to ALWB from Science Foundation Ireland (grant number 11/RFP.1/NES/3194). JASK is supported by a grant from the National Institute of Mental Health (MH080838), the Chaire d'Excellence Pierre de Fermat, and the Davimos Family Endowment for Excellence in Science.
Data were provided [in part] by the Human Connectome Project, WU-Minn Consortium (Principal Investigators: David Van Essen and Kamil Ugurbil; 1U54MH091657) funded by the 16 NIH Institutes and Centers that support the NIH Blueprint for Neuroscience Research; and by the McDonnell Center for Systems Neuroscience at Washington University.
Open Research
DATA AVAILABILITY STATEMENT
The data that support the findings of this study are available from the The Human Connectome Project (HCP) WU-Minn consortium at https://www.humanconnectome.org/, reference number 1200 Subjects Release of HCP image and behavioural data. These data were derived from the following resources available in the public domain: ConnectomeDB management platform (https://db.humanconnectome.org/app/template/Login.vm) and 1200 Subjects Data Release (https://www.humanconnectome.org/study/hcp-young-adult/document/1200-subjects-data-release).

