

Seabird bycatch vulnerability to pelagic longline fisheries: Ecological traits matter
Abstract
- Fisheries bycatch of seabirds presents a serious management problem, and relatively little is known about this problem. In the Western North Atlantic, the issue is complicated by the relatively low effort and coverage of the observer programme and the high uncertainty in species identification.
- The Western North Atlantic is home to many endemic and endangered seabird populations, and the impact of fishery-caused seabird bycatch has been of high interest, especially for those species with a low population size; however, species-specific bycatch estimates have been difficult. From 1992 to 2016, 158 seabirds were observed caught by the US Atlantic pelagic longline fleet; among them, only 80 were identified to species, 25 were identified to family, and the rest – mainly in the older records – were unidentified.
- In this study, ecological traits of seabirds were used to improve bycatch estimation and provide species-specific risk analysis to all the potentially affected seabird species in this region. Bayesian state–space modelling was used to accommodate the high level of uncertainty in the species identification process.
- Seabird bycatch risk was found to be highly dependent on population size. The group of large seabird species was estimated to be two times as vulnerable as the group of mid-to-small seabird species, scavenging and plunge-diving feeding modes were identified as imposing high bycatch risks, and spatial and temporal distribution patterns were also good indicators of bycatch risk. Based on these ecological traits, shearwaters, gulls, gannets, and petrels were identified to potentially suffer from high bycatch in this region. These species, especially those that have not been identified historically, deserve extra attention in the observer programme, and for the implementation of conservation measures of seabirds in this region.
1 INTRODUCTION
Fisheries bycatch presents a major threat to the biodiversity of seabirds around the world, with at least 160 000 seabirds removed by longline operations annually (Anderson et al., 2011; Hedd et al., 2016). Like many other animals (e.g. whales and turtles) incidentally caught by fishing gear, many seabirds are long-lived animals for which an increase in adult mortality can have an almost immediate effect on the population trend (Lewison et al., 2012). For example, fisheries bycatch at least partially contributed to the decline of some Pacific albatross populations (Croxall, Prince, Rothery, & Wood, 1998; Croxall, Rothery, Pickering, & Prince, 1990; Weimerskirch, Brothers, & Jouventin, 1997). The issue of seabird bycatch is a cooperative conservation and management problem that requires collaboration among managers, fishermen, and scientists. A successful seabird bycatch reduction programme should include three major tasks: (i) bycatch rate estimation; (ii) biological impact assessment; and (iii) the implementation of mitigation practices (Komoroske & Lewison, 2015; Lewison et al., 2012). The first task is fundamental, with the other two tasks being critically dependent upon the bycatch rate estimation.
Seabird bycatch is difficult to study because of the rarity of such events (Brothers, Duckworth, Safina, & Gilman, 2010; Li & Jiao, 2013). Seabirds are not targeted by fishing operations, and mitigation measures that reduce seabird bycatch are in place in some fisheries (Bull, 2007; Klaer, 2012; Klaer & Polacheck, 1998; Li, Browder, & Jiao, 2012; Watson, Epperly, Shah, & Foster, 2005). In contrast with drift-netting, which was banned in 1993 (Brothers, Cooper, & Lokkeborg, 1999), longlining has been regarded as an environmentally friendly fishing technique (Bjordal & Lokkeborg, 1996), but for a contrary view see Hinman (1998). More than 99% of observed US Atlantic pelagic longline (PLL) operations did not catch a seabird (Li & Jiao, 2013; Winter, Jiao, & Browder, 2011; Zhou & Jiao, 2017). Despite this relatively low rate, bycatch events could potentially be detrimental to many endemic and endangered seabird populations in the North Atlantic, such as the Bermuda petrel (Pterodroma cahow), for which the last census documented only 53 breeding pairs (International Union for Conservation of Nature (IUCN), 2017; no bycatch of this species has yet been recorded in US PLL operations). Thus, it is of conservation interest to assess the potential risk of fishing operations to these populations with a low abundance instead of just estimating a region-wide total seabird bycatch (Li & Jiao, 2013).
Seabird bycatch along the Western North Atlantic from the PLL fishery was used in this study. Currently, little is known about the seabird bycatch process because of the low quality and quantity of bycatch records from the PLL fishery, arising from the relatively low coverage of the observer programme, the rarity of seabird bycatch events, and the high uncertainty in species identification. Most fisheries around the globe either do not have an observer programme or only cover a small percentage of total effort (Anderson et al., 2011; Brothers et al., 1999). In the Western North Atlantic, the Pelagic Observer Program (POP) administered by the Southeast Fisheries Science Center has aimed for a coverage of 8% in recent years (Beerkircher, Cortes, & Shivji, 2002; Diaz, Beerkircher, & Restrepo, 2009). This coverage may be too low to reliably document the scale of seabird bycatch in the Atlantic, potentially impeding the implementation of any mitigation strategies. Bycatch estimation would improve with greater coverage, although such a strategy may be logistically and financially difficult. Furthermore, it would take time for the improved coverage to translate into a stronger conclusion. Studies to include both recent years with high observer effort and historical years with low observer coverage are common (Brothers et al., 1999; Li & Jiao, 2013). In the Western North Atlantic, POP data ranging back to 1992 was used in the analysis, with effort coverage ranging from 2 to 7% before 2004, and from 7 to 18% after 2004. The performance of morphology-based at-sea species identification employed in the POP was especially poor from 1997 to 2003, and resulted in just 16% of the total number of seabirds being identified to family level, with 34% left unidentified up to 2016. The situation has improved since 2004, with the initiation of a seabird identification training programme routinely scheduled along with other training activities each time a group of new observers is hired, and at other times (Beerkircher et al., 2002); however, as a result of the degradation of specimens during soaking and hauling time, positive identification cannot be obtained for some specimens, even with proper observer training and good photos.
In order to estimate seabird bycatch at the species or species-group level, the relationships between seabird bycatch likelihood and the ecological traits of seabirds were explored. Essentially, seabird bycatch is the consequence of interaction between the seabird community and fishing operations, mediated through the foraging behaviour and morphological traits of the birds, target-based fishing practices of individual vessels, and the temporal and spatial overlap of seabird populations with fishing vessels in areas of high productivity (Brothers et al., 1999). The ecological traits of the birds act as filters that assign seabirds to different levels of exposure to fishery operations (Figure 1). For example, surface scavengers, such as albatrosses, petrels, shearwaters, and gulls, that take baits from hooks are more vulnerable to longline fishing operations than deep-diving and non-scavenging species, such as penguins and cormorants, which are rarely caught on PLL (Brothers et al., 1999; Camphuysen et al., 1995). Deep-diving species are not spared, however, as two plunge-divers, the northern gannet (Morus bassanus) and the brown pelican (Pelecanus occidentalis), are also found as bycatch of US Atlantic PLL. Smaller species, such as terns and auklets, are rarely caught because PLL baits are too large to swallow (Brothers et al., 1999). No comprehensive analysis has previously been conducted to quantify the effects of these traits on seabird bycatch risk. This study started with the 48 seabird species that were initially recognized in the US North Atlantic fleet fishing footprint by Lee (2009), who synthesized morphological characteristics associated with fishing behaviour that make these species potentially differentially vulnerable to risk from the PLL fishing operations. The morphological and ecological traits of seabirds were integrated into a traditional fishery-centric bycatch analysis and investigated to improve their potential for seabird bycatch estimation. Then, the bycatch mortality rate of seabird species of conservation concern was assessed based on the morphological characteristics and ecological traits of the species.

2 METHODS
2.1 Seabird bycatch in the US Atlantic PLL fishery
The US Atlantic PLL fishery primarily targets yellowfin tuna (Thunnus albacares), bigeye tuna (Thunnus obesus), other tuna species, swordfish (Xiphias gladius), sharks, and dolphin fish (Coryphaena hippurus). Since the inception of the Atlantic observer programme in 1992, the observer effort in relation to the total fleet effort, in terms of number of sets deployed, has increased, averaging 7.03% of the total annual PLL fishing effort. The POP observed 18 913 longline sets from 1992 to 2016. Observers recorded not only bird bycatch, but also the bycatch of other incidental taxa, as well as the target species. Environmental information, spatial coordinates, target species, and set characteristics, such as number of hooks per set, were also recorded. The US Atlantic fleet operates in pelagic waters along the eastern seaboard from the Grand Banks off Newfoundland to the Florida Keys, and throughout the US Gulf of Mexico. Bird bycatch has occurred mainly in four of the 10 subregions identified for fishery management purposes: North-east Coastal (NEC), Mid-Atlantic Bight (MAB), South Atlantic Bight (SAB), and Gulf of Mexico (GOM) (Figure 2; Li & Jiao, 2013; Winter et al., 2011).

Both the Atlantic pelagic observer and PLL logbook data for 1992–2016 were provided by the Fisheries Statistics Division of the Southeast Fisheries Science Center of the National Marine Fisheries Service, Miami, Florida. Data consistency was checked between the two types of data sets. Records with obviously erroneous or missing necessary information, such as geographic coordinates or number of hooks, were excluded from the analysis. A total of 18 726 longline sets with 83 sets of positive bycatches from the observer data were used for model fitting, and a total of 266 324 longline sets from the fishery logbook data were used for prediction.
2.2 Ecological traits of seabirds
Body size, feeding behaviour, and spatial and temporal distribution were extracted from the literature (Kaufman, 2001; Lee, 2009; Rodewald, 2015), and seabird species composition data were extracted from the at-sea survey data in the United States Geological Survey (USGS) Avian Compendium Database (O'Connell et al., 2011). A total of 48 seabird species distributed in the Western North Atlantic were identified as potential species susceptible to PLL operations (Lee, 2009). The inclusion of a species was based on either historical identified bycatch in the Atlantic or the known diet and foraging behaviour of the species. Seabird species profiles compiled by Lee (2009) for POP training were used as the main source of references of body size and spatial distribution. Noted ecological traits were checked against Kaufman (2001) and Rodewald (2015).
All seabirds from the fishing region were classified into three size groups based on body length to help the observers identify them (Lee, 2009). Of the 48 species vulnerable to longline operations, 10 species have been historically identified in the POP bycatch records, including four species in the large species category (brown pelican, northern gannet, great black-backed gull (Larus marinus), and herring gull (Larus argentatus)), five species in the mid-sized species category (northern fulmar (Fulmarus glacialis), Cory's shearwater (Calonectria diomedea), greater shearwater (Ardenna gravis), parasitic jaeger (Stercorarius parasiticus), and the laughing gull (Larus atricilla)), and one species in the small species category (Wilson's storm petrel (Oceanites oceanicus)) (Table S1). With the limited number of observed ‘small’ species (only one recorded specimen), the last two categories were combined as the mid-to-small species category.
Seabirds employ a diverse set of feeding tactics, and these feeding tactics can be categorized into surface feeding, plunge-dive feeding, pursuit diving, and scavenging (Haney & Stone, 1988). The main mode(s) of feeding tactics of each seabird species were obtained from Kaufman (2001) and Rodewald (2015). Some species primarily employ just one feeding mode: for example, the northern fulmar is primarily a surface feeder but may also plunge into the water and dive (Kaufman, 2001). Only the primary mode of feeding tactic was recorded for these species. Other species may primarily employ multiple feeding modes: for example, Cory's shearwater forage by plunge diving from near the surface, and also take items from the surface of the water (Kaufman, 2001). Multiple feeding modes were recorded for these species.
The spatial and temporal distributions of these seabirds were also included in the analysis (Kinlan, Winship, White, & Christensen, 2016; Lee, 2009; O'Connell et al., 2011). Two levels of spatial variation (expected and unexpected to occur in 11 statistical zones in the Western North Atlantic) and two levels of temporal variation (common and uncommon) were used independently of other ecological traits. Seabirds endemic to the Western North Atlantic were recorded as ‘common’ in all seasons. When information in the literature was insufficient to distinguish seasonal differences, ‘unknown’ was recorded for the respective species and precautionarily treated as ‘common’ in the modelling process.
The composition of seabirds vulnerable to the Atlantic PLL fishery was treated as an index of the background seabird abundance in this study and was used to test the density-dependent effect on seabird bycatch. The USGS Avian Compendium has a spatial coverage along the Atlantic coast of the USA from Florida to Maine (Kinlan et al., 2016; O'Connell et al., 2011). As a result of limited coverage for some years, data from all surveys in the database were pooled across time to generate a single composition table. Four of the 48 species identified as potentially susceptible to PLL operations were not positively sighted in any of the surveys from the database; these species include soft-plumaged petrel (Pterodroma mollis), red-footed booby (Sula sula), ivory gull (Pagophila eburnean), and Ross's gull (Rhodostethia rosea). For these species, setting the number of observed individuals to zero would agree with the observed data but would eliminate the bycatch possibility of these species; on the other hand, setting them to one would contradict the observed data. Without further information on the relative density of these species, non-informative priors (uniform between zero and one) can be assumed for these values, and a Monte Carlo scheme can then be used to integrate over those non-informative priors to assess the bycatch uncertainty associated with these species; however, this approach is computationally intensive. To simplify computation, only the means of these priors (a one-half observation) were used for these species to represent the average case, and a further analysis of the posterior bycatch estimates of these species indicated that a full Monte Carlo scheme is not needed (see results section for further information). All other species have integer values equal to or larger than one in the composition table (Table S1).
2.3 Ecological hypotheses on the fishery interaction of seabirds with PLL gear
Six ecological hypotheses were formulated based on the differential interaction among seabird species and the spatial and temporal overlap of seabird populations with the US PLL fleet. The null hypothesis (H0) postulated that there were no density effects as a result of the differential distribution in species identity, space, and time, and no morphological effects in the fishery interaction of seabirds among all species. It forms the baseline scenario against which other hypotheses can be compared. In the null hypothesis, each species was assumed to have a unit relative abundance, i.e. Nk = 1, where k = 1…48 is the index for species names, and the relative abundance of each species is evenly distributed among seasons and spatial locations.
The rest of the hypotheses were formed by using the ecological traits of seabirds to adjust the level of exposure of underlying seabird populations to PLL operations. First, we tested whether seabird density alone is a good indicator of differential bycatch risk among species (H1). In H1, the species composition from the Avian Compendium database was used for the underlying relative species abundances, i.e. Nk = nk, where nk is the number of observed individuals of species k in the Avian Compendium database. Again, the relative abundance of each species is assumed to be evenly distributed among seasons and spatial locations.
Then, the effect of species size (i.e. size group) on bycatch risk (H2) was tested. In addition to the species-specific relative abundances used in H1, H2 incorporated varying susceptibility with respect to the normal size of the species. The proportion of seabirds from species k that is susceptible to PLL bycatch is Nk = nk × susk, where susk measures the susceptibility of species k on a scale from zero to one, and H1 is a special case of H2 where all the species have the same susceptibility. Two levels of susceptibility were used, corresponding to the two size categories (large species and mid-to-small species). As the susceptibility used here measures the relative risk of bycatch, the two levels of susceptibility cannot be estimated independently, and so the susceptibility coefficient of the mid-to-small species was fixed to 0.01, and the susceptibility of the large species was given a non-informative prior Beta(1,1) uniform over zero and one. Setting smid to small = 0.01 bounded the upper limit of the susceptibility of large species, slarge, to be at most 100 times the value for smid to small, but the lower limit was left unbounded. Results demonstrated that the pre-set level did not constrain the estimation of slarge.
Next, the effect of feeding tactics on bycatch risk given size differences among species (H3) was tested. Four modes of feeding tactics were explored, i.e. surface feeding, plunge diving, pursuit diving, and scavenging. In H3, the relative bycatch risk conferred by each of these feeding tactics was estimated. The lack of a feeding mode of a species was assigned a fixed 1% reference bycatch risk, and each mode of feeding tactic was given a priori bycatch risk of U(0,1): for example, a smaller than 1% value indicates that the given feeding mode reduces bycatch risk, and a larger than 1% value indicates that the given feeding mode increases bycatch risk. The reference bycatch risk level used here imposes the upper limit of the bycatch risk of any feeding tactic to be at most 100 times as risky as the reference level. It is further assumed that there is no interaction among modes of feeding behaviours. Thus, in H3 the proportion of seabirds from species k susceptible to PLL bycatch is Nk = nk × susk × feedingk, with feedingk = SFk × PLk × PSk × SCk, where SF is given a prior of Beta(1,1) if the given species is a surface feeder (SF), and is set to the reference level (1%) if surface feeding is not among its main feeding tactics, and the parameters for plunge diving (PL), pursuit diving (PS), and scavenging (SC) are set in the same manner.
Finally, the effect of spatial location and season on bycatch risk through their impact on the distribution of the relative abundances and composition of the seabird community was tested. Two additional hypotheses were formulated based on H3. The temporal hypothesis (H4) postulates that seasonal migratory patterns of seabirds modulate their exposure to PLL bycatch. Two levels of exposure were used to measure the changes in abundance levels among seasons, based on historical patterns in the seasonal occurrences (common and uncommon) retrieved from the literature indicated previously. As the two levels of temporal exposure cannot be independently estimated, the temporal exposure coefficient for ‘common’ seasons was fixed to 1, and the exposure coefficient for ‘uncommon’ seasons was given an uninformative prior of Beta(1,1), uniform between zero and one. Under H4, the relative abundance of seabird species k in season l susceptible to bycatch is Nk,l = nk × susk × feedingk × tk,l, where tk,l is the temporal exposure coefficient of species k in season l. The underlying frequency of species identity α in season l is
 , where the first summation is over all of the species under species identity α.
, where the first summation is over all of the species under species identity α.

if area k is one of the expected occurrence areas, and 0 otherwise, where ak is the number of areas where species k is expected to occur. The underlying frequency of species identity α in season l and area m is
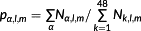 , where the first summation is over all of the species under species identity α.
, where the first summation is over all of the species under species identity α.
2.4 Binary process

2.5 Counting process


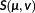 is a normalizing constant, v ≥ 0 is the shape parameter, and μ > 0 is a centring parameter of the CMP distribution, with the assumption that log(μ) has a linear relationship with the covariates. The modification of the CMP distribution involves mapping the singleton catch event to the sum of probabilities of outcomes of less than k + 1 from the original distribution, and the probability of outcomes of larger than k + 1 is mapped to the probability of an outcome of n + k – 1 from the original distribution (Zhou & Jiao, 2017). With the small sample size of positive bycatch records, only one covariate, the number of hooks, was used in the counting process submodel. The number of hooks deployed is a frequently used measure of PLL fishing effort in bycatch studies (Brothers et al., 1999; Gilman, Chaloupka, Peschon, & Ellgen, 2016; Li, Jiao, & Browder, 2016).
is a normalizing constant, v ≥ 0 is the shape parameter, and μ > 0 is a centring parameter of the CMP distribution, with the assumption that log(μ) has a linear relationship with the covariates. The modification of the CMP distribution involves mapping the singleton catch event to the sum of probabilities of outcomes of less than k + 1 from the original distribution, and the probability of outcomes of larger than k + 1 is mapped to the probability of an outcome of n + k – 1 from the original distribution (Zhou & Jiao, 2017). With the small sample size of positive bycatch records, only one covariate, the number of hooks, was used in the counting process submodel. The number of hooks deployed is a frequently used measure of PLL fishing effort in bycatch studies (Brothers et al., 1999; Gilman, Chaloupka, Peschon, & Ellgen, 2016; Li, Jiao, & Browder, 2016).2.6 Species composition
Difficulty in positively identifying a specimen may come from either a lack of proper observer training or the degradation of a specimen during the hauling process. The first issue can be eliminated through improved training, whereas the second issue is likely to remain with the current morphology-based species identification protocol of the POP. To accommodate the uncertainty in classification to species, state–space modelling techniques were used in this analysis: the 48 seabird species susceptible to PLL fishing were modelled as state variables, and only a select number of species could be directly observed; the rest of the species were only observable at either the family or class level. The 12 fully or partially observable species or species groups are referred to as ‘species identities’ later in the text. Each of the first 10 identities comprised a single species, i.e. northern fulmar, Cory's shearwater, greater shearwater, Wilson's storm petrel, brown pelican, northern gannet, parasitic jaeger, great black-backed gull, American herring gull, and laughing gull. The 11th and 12th identities include multiple species, i.e. the shearwater group, comprising six species out of the 48, including those identities mentioned above, and the gull Laridae family, comprising 15 species out of the 48, including those mentioned above.

2.7 Bayesian approach
A Bayesian approach was used for parameter estimation. Normal priors with mean zero and variance 10 were used on rescaled covariates (zero mean and unit variance). For distribution-specific shape parameters, wide uniform priors were used. To simulate Markov chain Monte Carlo (MCMC) samples from the posterior distribution, jags 4.0 (Plummer, 2003) was used in the statistical program r 3.2.5 (R Development Core Team, 2016).

 is the posterior mean of the deviance, and pD is an estimate of the effective number of parameters in the model based on the algorithm proposed by Plummer (2002). A smaller DIC value indicates a better model. DIC was calculated conditionally on the estimated year effect because of the goal to predict total bycatch for each of those years.
is the posterior mean of the deviance, and pD is an estimate of the effective number of parameters in the model based on the algorithm proposed by Plummer (2002). A smaller DIC value indicates a better model. DIC was calculated conditionally on the estimated year effect because of the goal to predict total bycatch for each of those years.3 RESULTS
Based on DIC, the full model (H5) turned out to be the best-supported model (Table 1). H5 outperformed the null hypothesis H0 by a reduction of 329.2 points in DIC, indicating the utility of using ecological traits to estimate bycatch risk. Seabird relative abundance, size, feeding behaviour, seasonal emigration patterns, and large-scale spatial distribution patterns reduced model uncertainty in predicting seabird bycatch risk. Among the factors that improved bycatch risk prediction, the inclusion of seabird relative abundances based on the Avian Compendium introduced the largest decrease in DIC (Table S2). Additionally, Watanabe–Akaike information criterion (WAIC) values were calculated based on the posterior samples and the model selection results agree with those based on DIC (Table S2).
| Hypothesis | Description | ΔDIC |
|---|---|---|
| H0 | Null hypothesis: no ecological effects | 329.2 |
| H1 | Density hypothesis: H0 + relative abundance | 70.6 |
| H2 | Size-selective hypothesis: H1 + size-related susceptibility | 46.1 |
| H3 | Feeding-tactics hypothesis: H2 + feeding tactics | 25.0 |
| H4 | Temporal hypothesis: H3 + seasonal migration | 5.5 |
| H5 | Spatial–temporal hypothesis: H4 + spatial distribution | 0.0 |
In addition, the body size of the seabird species is a strong indicator of PLL bycatch risk. The inclusion of size selectivity further improved model performance. Based on the selected model, the distribution of the posterior estimates of slarge clearly indicates that large species are substantially more vulnerable to PLL operations than smaller species (Figure 3). The bycatch susceptibility of large species, such as the northern gannet, is 2.03 times that of the mid-to-small species, such as Cory's shearwater, with a 95% credible interval of 1.22–2.97 times.

Species with scavenging and plunge-diving feeding modes were identified to have higher risks of bycatch while interacting with PLL operations. Posterior estimates of vulnerability parameters of scavenging and plunge-diving modes significantly moved away from their prior distribution (uniform from 0 to 1 in this case), suggesting supporting evidence in the dataset (Figure 4). Furthermore, 99.33% of the posterior estimates of the vulnerability of the scavenging feeders were above the reference level, and 100% of the posterior estimates of the vulnerability of the plunge-diving mode were above the reference level, indicating significantly higher risks for these two feeding modes. The scavenging and plunge-diving species were estimated to increase the bycatch risk by on average 2.08 and 4.16 times, respectively, in the fishery interaction of seabirds. On the other hand, the posterior estimates for the vulnerability of species that demonstrate surface feeding and species that demonstrate pursuit diving resemble their prior distributions, suggesting little evidence in the data to infer the vulnerability of species with these feeding modes.

The seasonal migratory pattern of seabirds was also found to be a strong indicator of their bycatch risk. Of the 80 seabird specimens identified to species level, four were caught in uncommon seasons, with the rest of them caught in common seasons. Seabirds identified at the family level and unidentified seabirds also contributed to the estimation of the seasonal exposure to bycatch risk. Adding seasonal migratory pattern into the model further increased the model performance. Based on the selected model, the exposure of seabirds in their respective uncommon seasons is on average 17.65% of their exposure during the common seasons. The upper 95% percentile of the posterior estimate of the exposure in uncommon seasons, tuncommon, is 34.23%, substantially smaller than the exposure in common seasons (Figure 5).

The incorporation of the large-scale spatial distribution of seabirds gave the model a further boost in performance. Three characteristics of the distribution, overlain on the background of observed effort, are especially important (Figure 2): (i) almost all observed bycatch events occurred in five regions, which were, from east to west, NED, NEC, MAB, SAB, and GOM; (ii) the bycatch events in these high-risk regions were not uniformly distributed among areas or within each area; and (iii) the observed bycatch events fell within a narrow band stretching from north-east to south-west, with the exception of just one observation from TUN, suggesting that the underlying structuring force of the observed regional pattern may be the proximity of the fishing locations to the mainland. The coarse-scale spatial distribution information used in this study was helpful in revealing the pattern of bycatch risk at the scale of the defined PLL fishing regions.
The total species-specific bycatch of the US Atlantic PLL fleet from 1992 to 2016 was projected from the logbook effort data based on parameter estimates of the selected model (Table 2). The posterior estimate of the total number of seabird bycatch from 1992 to 2016 was on average 3194 seabirds, with a 95% credible interval of 2292–4297 seabirds. Out of the 10 historically identified seabird species, the herring gull (1491) was the most frequently caught species, followed by the greater shearwater (1035). On average, 6.58% of the shearwater bycatch does not belong to the recorded bycatch species of this family, and 1.58% of the gull bycatch does not belong to the gull species recorded as bycatch. Black-capped petrel (Pterodroma hasitata; 24), sooty shearwater (Ardenna grisea; 67), and Audubon's shearwater (Puffinus lherminieri; 44) were predicted to face high bycatch risk, although they have not been historically identified in the POP records. The projected impact of the US PLL fleet on the Bermuda petrel (0), which is a rare species, was found to be negligible. The four species assigned a one-half observation in the composition table were absent from the projected bycatch (Table 2). This indicated that, on average, these species were not subject to bycatch risks based on the current assessment model, and that the current treatment of missing observations was sufficient. Thus, the full Monte Carlo scheme was not further pursued. For the complete list of projected bycatches for all 48 seabird species included in the model, see Table 2.
| Family | Species name | Mean | Lower 2.5 percentile | Upper 2.5 percentile |
|---|---|---|---|---|
| Loons Gaviidae | Common loon (Gavia immer) | 1 | 0 | 5 |
| Shearwaters Procelleriidae | Northern fulmar (Fulmarus glacialis) | 6 | 1 | 15 |
| Bermuda petrel (Pterodroma cahow) | 0 | 0 | 0 | |
| Black-capped petrel (Pterodroma hasitata) | 24 | 8 | 48 | |
| Herald petrel (Pterodroma arminjoniana) | 0 | 0 | 0 | |
| Soft-plumaged petrel (Pterodroma mollis) | 0 | 0 | 0 | |
| Bulwer's petrel (Bulweria bulweria) | 0 | 0 | 0 | |
| Cory's shearwater (Calonectris diomedea) | 67 | 33 | 116 | |
| Greater shearwater (Ardenna gravis) | 1035 | 656 | 1503 | |
| Sooty shearwater (Puffinus griseus) | 31 | 14 | 58 | |
| Manx shearwater (Puffinus puffinus) | 2 | 0 | 6 | |
| Audubon's shearwater (Puffinus lherminieri) | 44 | 20 | 78 | |
| Little shearwater (Puffinus assimilis) | 0 | 0 | 1 | |
| Storm petrels Hydrobatidae | Wilson's storm petrel (Oceanites oceanicus) | 91 | 37 | 173 |
| Leach's storm petrel (Oceanodroma leucorhoa) | 6 | 1 | 14 | |
| Band-rumped storm petrel (Oceanodroma castro) | 1 | 0 | 3 | |
| White-faced storm petrel (Pelagodroma marina) | 0 | 0 | 0 | |
| Tropicbirds Phethontidae | Red-billed tropicbird (Phaethon aethereus) | 0 | 0 | 1 |
| White-tailed tropicbird (Phaethon lepturus) | 0 | 0 | 2 | |
| Pelicans Pelecanidae | Brown pelican (Pelecanus occidentalis) | 3 | 0 | 8 |
| Gannets and boobies Sulidae | Northern gannet (Sula bassana) | 89 | 44 | 161 |
| Masked booby (Sula dactylatra) | 6 | 1 | 13 | |
| Brown booby (Sula leucogaster) | 0 | 0 | 0 | |
| Red-footed booby (Sula sula) | 0 | 0 | 0 | |
| Auks and puffins Alcidae | Atlantic puffin (Fratercula arctica) | 0 | 0 | 2 |
| Razorbill (Alca torda) | 1 | 0 | 3 | |
| Common murre (Uria aaige) | 0 | 0 | 1 | |
| Thick-billed murre (Uria lomvia) | 0 | 0 | 2 | |
| Jaegers and skuas Stercorariidae | Parasitic jaeger (Stercorarius parasiticus) | 1 | 0 | 3 |
| Long-tailed jaeger (Stercorarius longicaudus) | 0 | 0 | 1 | |
| Pomarine jaeger (Stercorarius pomarinus) | 7 | 1 | 15 | |
| Great skua (Catharacta skua) | 1 | 0 | 4 | |
| South polar skua (Catharacta maccormicki) | 1 | 0 | 3 | |
| Gulls Laridae | Great black-backed gull (Larus marinus) | 235 | 116 | 406 |
| Glaucous gull (Larus hyperboreus) | 0 | 0 | 1 | |
| Iceland gull (Larus glaucoides) | 0 | 0 | 1 | |
| Herring gull (Larus argentatus) | 1491 | 997 | 2121 | |
| Lesser black-backed gull (Larus fuscus) | 0 | 0 | 1 | |
| Ring-billed gull (Larus delawarensis) | 16 | 6 | 28 | |
| Ivory gull (Pagophila eburnea) | 0 | 0 | 0 | |
| Black-legged kittiwake (Rissa tridactyla) | 9 | 2 | 19 | |
| Laughing gull (Larus atricilla) | 19 | 6 | 39 | |
| Common black-headed gull (Larus ridibundus) | 0 | 0 | 0 | |
| Franklin's gull (Larus pipixcan) | 0 | 0 | 0 | |
| Bonaparte's gull (Larus philadelphia) | 3 | 0 | 9 | |
| Ross’ gull (Rhodostethia rosea) | 0 | 0 | 0 | |
| Sabine's gull (Xema sabini) | 0 | 0 | 0 | |
| Little gull (Larus minutus) | 0 | 0 | 0 |
4 DISCUSSION
This study documents the first effort to incorporate the ecological traits of seabird species into their bycatch risk analysis to provide a species-specific total bycatch estimate in the Western North Atlantic. Seabird species susceptible to PLL operations have different bycatch risks with respect to their ecological traits. This study quantified the effects of factors including relative abundance, size (total length), feeding behaviour, and spatial and temporal migratory pattern on seabird bycatch in the Atlantic PLL fishery, and estimated the total species-specific bycatch in this region. The usage of this ecological approach is not restricted to the US Atlantic PLL fisheries, and it can be applied to other fisheries to provide a species-specific bycatch risk assessment. Further study to explore the distribution of seabird bycatch risks on a finer spatial scale would further facilitate efforts to mitigate seabird bycatch reduction.
Relative seabird density was found to be an important indicator of PLL seabird bycatch. This result agrees with other studies where a positive relationship was found between bycatch rates and local seabird abundances (Gilman et al., 2012, 2016; Gilman, Boggs, & Brothers, 2003; Reid & Sullivan, 2004; Ryan, Keith, & Kroese, 2002). The use of the Avian Compendium database in this study is relatively crude in that only the overall pattern of species composition is used to predict bycatch risk. Nevertheless, the overall species composition proved to be the most important factor in predicting bycatch risk among the five factors applied in this study. The model with species composition included performed much better, not only than the model without ecological traits (null hypothesis H0) but also than the model using the relative abundance of each bird species (data from Lee, 2009) as a density-dependent factor in a preliminary analysis, which were seen through the large reductions in DIC and WAIC. This result encourages future seabird bycatch studies to use regional or local relative abundance information from at-sea surveys when such information is available. Meanwhile, finer grained information on the spatial and temporal distribution patterns of seabirds in the Western North Atlantic could be extracted for some seabird species from this database (but not all of them) (Kinlan et al., 2016). Such detailed spatial information can be used to further improve bycatch analysis, and this topic deserves further investigation.
Seabird bycatch from the PLL fishery is a highly size-selective process. Small species are rarely observed being captured by longliners (Gilman et al., 2016; Zhou & Jiao, 2017), and it is hypothesized that small seabird species are seldom caught because they are unable to swallow the relatively large baits (Brothers et al., 1999). This is the first time that the size selectivity of the PLL seabird bycatch has been quantified based on observation data. The analysis showed that the large species group was around twice as vulnerable as the mid-to-small species group. As k-selected animals, seabird populations are most sensitive to changes in adult mortality (Gilman et al., 2016; Lewison et al., 2012), and a highly skewed mortality against larger adult birds implies that the biological impact of PLL bycatch on seabird populations could be more severe than previously thought (Gianuca, Phillips, Townley, & Votier, 2017).
Scavenging and plunge diving were identified as high-risk feeding tactics that would increase the bycatch probability of seabirds when interacting with PLL operations. There is no doubt that the scavenging and diving behaviour of some seabird species cause greater exposure to PLL operations (Brothers et al., 1999). The interaction of seabirds and PLL operations typically occurs at the line-setting stage, when the baited hooks are near the surface of the sea, and also at the hauling stage when seabirds try to take the remaining baited hooks (Huin & Croxall, 1996). This study gives ecological evidence that bycatch reduction strategies that aim to reduce the availability of the baited hooks near the sea surface, such as the use of weighted hooks and the underwater setting of hooks (Bull, 2007; Gilman et al., 2003; Ryan & Watkins, 2002), would be particularly effective in mitigating seabird bycatch in this region.
Seabirds found in the Western North Atlantic exhibit a diverse set of temporal migratory patterns, and these patterns are good indicators of their bycatch risk. Many seabird species are present throughout the year, e.g. the black-capped petrel (Golovkin, 1984) and the Atlantic puffin (Fratercula arctica) (Brown, Brown, & Nettleship, 1989). Some migrants appear in the Western North Atlantic only seasonally: Sabine's gull (Xema sabini) from eastern Canada breed in the Arctic and mainly migrate eastwards across the North Atlantic, and then south to winter mostly in South Africa (Kushlan et al., 2002); the common loon (Gavia immer) nests in fresh water but can be expected at sea in winter (Haney, 1990). Meanwhile, some obligate trans-equatorial migrants, such as Cory's shearwater, the greater shearwater, and the sooty shearwater, move from the southern hemisphere to the Western North Atlantic in summer (Croxall, Evans, & Schreiber, 1984). Some species are only present occasionally in the Western North Atlantic as vagrants, such as the lesser black-backed gull (Larus fuscus) (Golovkin, 1984). Factors other than migration may also induce seasonal variations in bycatch risk, e.g. the chick-rearing needs of Laysan and black-footed albatross may have increased their bycatch risk during the breeding season (Fernández, Anderson, Sievert, & Huyvaert, 2001; Gilman et al., 2016; Hyrenbach, Fernández, & Anderson, 2002). As the breeding ground of many of the bycatch species included in this assessment does not overlap with the study area (Western North Atlantic), breeding season alone would not be a good indicator of the presence or absence of a seabird species in this region. On the other hand, reproductive season could be included in future bycatch assessment models to investigate its effects on bycatch species that breed in the Western North Atlantic.
Most of the difficulties associated with studying seabird bycatch come from the low number of direct observations. This study revealed that ecological traits are important factors in determining a seabird's bycatch vulnerability to the PLL fishery, and harnessed the ecological traits of seabirds to improve the task of bycatch estimation by providing species-specific vulnerability and bycatch estimates. Another approach to advance seabird bycatch analysis is to improve the quality of the data collection process: for example, the seabird identification training programme organized by J. Browder as part of the US National Oceanic and Atmospheric Administration (NOAA) Fisheries’ National Seabird Program has dramatically reduced the percentage of unidentified seabird bycatch in recent years. Based on ecological traits, this study identified seabird species that potentially suffer high bycatch risk. The species that have not been identified in the historical records need to be given extra attention in the observer training programme. It is acknowledged that species identification in the field will always have uncertainty given the unavoidable degradation of the physical condition of a sizable portion of seabird bycatch, especially those caught during the line-setting phase (Brothers et al., 2010). A combination of these strategies would promote the understanding of the bycatch process and help the mitigation efforts of seabird bycatch from the PLL fishery in the Western North Atlantic. In summary, using an ecological approach, this study provided a regional assessment of the species-specific bycatch risks for all of the seabird species vulnerable to longline operations in the Western North Atlantic; the proposed ecological approach operates on seabird ecology, historical bycatch, and logbook data, and does not require additional field sampling, making it widely applicable to other fisheries for a full species-specific bycatch risk assessment.
ACKNOWLEDGEMENTS
The US Atlantic PLL logbook data and POP data used in this study were provided by the Fisheries Statistics Division of the Southeast Fisheries Science Center, NOAA National Marine Fisheries Service, Miami, Florida. We thank Q. Ma for checking the ecological traits of seabirds against the literature. This research was supported by a contract for Modeling Pelagic Longline Seabird Bycatch awarded to Y. Jiao by the NOAA, National Marine Fisheries Service Southeast Fisheries Science Center (SEFSC), under a grant from the NOAA Fisheries’ National Seabird Program.

