Natural selection In Utero: Evidence from the great East Japan earthquake
Abstract
Objectives
Controversy remains over whether declines in male births reported after population stressors result from either or both reduced conception of males or increased selection in utero against male fetuses. We use monthly birth cohorts to determine if Japanese male births following the Great East Japan Earthquake of 2011 fell below levels expected from female births and from history (i.e., autocorrelation) among cohorts exposed to the Earthquake at or after conception.
Methods
We apply interrupted time-series methods to 69 months (i.e., April, 2006 through December, 2011) of birth data from the most and least affected prefectures as well as from the remainder of Japan. We estimate expected male births from female births and from autocorrelation.
Results
Findings varied by distance from the greatest damage but suggest sensitive periods both early and late in gestation when population stressors may induce selection against males in utero. Support for reduced conception of males appeared only in the prefectures most damaged by the Earthquake.
Conclusions
Results align with the claim that natural selection has conserved mechanisms that reduce the odds of a male live birth during stressful times by reducing the conception of males and by increasing the rate of spontaneous abortion among male fetuses. Am. J. Hum. Biol. 25:555–559, 2013. © 2013 Wiley Periodicals, Inc.
The argument that natural selection has conserved mechanisms by which human females spontaneously abort their least fit fetuses, particularly small males, enjoys wide attention if not acceptance (Trivers and Willard, 1973; Stearns, 1987; Forbes, 1997; Møller, 1997; Wells, 2000; Baird, 2009; Catalano et al., 2012b). This attention arises, at least in part, from the parsimony with which the argument explains, post hoc, the facts that at least 70% of human fertilizations spontaneously abort before birth or die during birth and that observable abortuses exhibit higher rates of abnormalities than do live born infants (Kline and Stein, 1987; Boklage, 1990). The argument also explains, again post hoc, the excess of small males among spontaneously aborted fetuses without abnormalities (Byrne and Warburton, 1987; Wells, 2000; Baird, 2009). Natural selection would conserve any mutation that spontaneously aborted the gestation of small males because, if born, they die more frequently before reproductive age than do larger males and much more frequently than do females of the same size or larger (Lummaa, 2001; Kristensen et al., 2007). Small males that survive to reproductive age, moreover, have less success in mating than do larger males (Sear, 2010). This relatively low fitness persists despite the fact that mothers routinely invest more of their resources in sons than in daughters (Clutton-Brock, 1991; Rickard et al., 2007; Powe et al., 2010).
The literature argues that selection in utero becomes less tolerant of impediments to fitness when population stressors trigger the maternal stress response (Catalano et al., 2012b). This argument assumes the common definition of stress as the physiological circumstance that arises when exogenous stressors approach or exceed the capacity of an organism to adapt. Selection in utero would be less tolerant among stressed women supposedly because the stressor may not only deplete maternal capacity to invest in infants but also increase the need for maternal investment in them.
Speculation on when in gestation an exogenous stressor will most likely trigger selection against less fit males arises from what little we know of spontaneous abortion (Catalano et al., 2012b). Fetal loss early in gestation remains difficult to observe, but research suggests that the highest rates for all fetuses appear in the first trimester and then decline until approximately the 16th week of gestation when the risk becomes stable (Byrne and Warburton, 1987; Goldhaber and Fireman, 1991).
Abortuses in the first trimester exhibit high rates of chromosomal abnormalities and morphological anomalies that profoundly threaten fitness regardless of fetal sex or maternal resources (Kline and Stein, 1987). These spontaneous abortions would, therefore, likely occur in benign as well as stressful environments.
The risk of spontaneous abortion among chromosomally normal fetuses also peaks, although less dramatically than among abnormal fetuses, around the 12th week and also declines to a 16th week, though higher, stable period (Kline and Stein, 1987). Unlike the risk among fetuses with identifiable abnormalities, the risk exhibits a second peak around 18 weeks and then declines slowly until the 24th week when it drops again (Kline and Stein, 1987). Small males predominate among the 16th to 24th week normal abortuses (Byrne and Warburton, 1987; Catalano et al., 2012a). This pattern leads to the speculation that while environmental stressors may induce selection against less fit males throughout pregnancy, its effects should be most apparent in the 16th through 24th weeks of gestation.
A rival to natural selection in utero as an explanation of reduced male births following population stressors remains because ambient stressors may reduce the conception of males by changing the hormonal environment at coitus (James, 2006, 2008). Although both explanations predict fewer than expected male births following population stressors, they imply different lengths of elapsed time between such stressors and their observable effect on male births. Reduced conception of males predicts declines 8–10 months after a stressor whereas selection in utero predicts a much quicker response. Much of the research, with the exception of the literature studying earthquakes, does not establish when in gestation a stressor predicts fewer male births.
The literature studying the likelihood of male births after earthquakes has used monthly data to estimate the delay between stressor and response. The findings, however, do not converge. One study reports no association (Tan et al., 2009). Another (Torche and Kleinhaus, 2012) reports a 3-month delay consistent with selection in utero. And 3 find support for reduced conception of males in that the ratio of male to female live births (i.e., secondary sex ratio) reportedly fell below expectations 9–11 months after the quakes (Fukuda et al., 1998; Saadat, 2008; D'Alfonso et al., 2012). Among these five studies, however, only that supporting selection in utero (i.e., Torche and Kleinhaus, 2012) used data of sufficiently understood provenance for inclusion in the Human Mortality Database (University of California Berkeley, 2012) and controlled for autocorrelation as well as secondary sex ratio changes in comparison populations. The study reporting no association (i.e., Tan et al., 2009) and two of those supporting reduced conception of males (i.e., Saadat, 2008; D'Alfonso et al., 2012) compare pre-earthquake and postearthquake births estimated from hospital records or from sources with unknown accuracy and coverage. The third study (i.e., Fukuda et al., 1998) supporting reduced conception of males used validated vital statistics, but does not control for autocorrelation in the data or for changes in male births that occur in similar but unexposed populations.
We contribute to the literature by assessing how long, if at all, after exposure to an unquestionably acute population stressor a well-monitored human population yielded lower than expected male births. More specifically, we measure the delay between the Great East Japan Earthquake and decline in male births across populations varying in distance from the area with most damage. We estimate the expected number of male births from the number of female births and from autocorrelation.
On March 11, 2011, a massive earthquake and subsequent Tsunami struck East Japan. It was the most severe earthquake ever in Japan and one of the most powerful earthquakes recorded anywhere in the last century (Geller, 2011). As of July 2012, The National Police Agency of Japan confirmed 15,867 deaths, 2,904 missing, and 6,109 injured (National Police Agency of Japan, 2012). Nearly all (i.e., 99.6%) the deaths occurred in three prefectures (administrative regions in Japan): Iwate, Miyagi, and Fukushima. While the earthquake and Tsunami induced numerous sequelae (e.g., nuclear plant disasters, and economic loss) affecting all Japan, the prefectures on the island of Kyushu suffered the least seismic activity and least damage (Japan Meteorological Agency, 2011; National Police Agency of Japan, 2012).
The argument for selection in utero assumes that the maternal stress response, if triggered during the 16th–24th week of gestation, will raise the standard not only for size but also for other signals of fitness required to continue the gestation of males (Catalano et al., 2012b). Over 80 percent of live births in Japan, as elsewhere, occur in the 36th through 40th week of gestation (Uehara et al., 2011). Based on this distribution, the effect of an exogenous shock on the secondary sex ratio should appear in birth data 12 weeks, or roughly 3 months, after the shock. This early effect would appear in the cohort exposed at the 24th week of gestation (i.e., latest week in the critical period), and born at the 36th week (i.e., earliest week of probable birth period). The last effect would appear 24 weeks, or roughly 6 months, after the shock in the cohort exposed at the 16th week of gestation (i.e., earliest week in the critical period) and born the 40th week (i.e., latest week of the probable birth period). The “window” in which we would expect to find an effect of the March 11 Great East Japan Earthquake, therefore, would extend from roughly June through August 2011. If, conversely, the Earthquake reduced the conception of males, we would expect to find an effect in November or December.
Our test asks whether the cohorts of infants exposed to the Earthquake at conception, or either early or late in gestation, included fewer than expected males. We also ask whether any differences between observed and expected males varied by distance from the most affected prefectures. We derive expected male births from female births and from autocorrelation.
MATERIALS AND METHODS
Data
We retrieved the monthly number of male and female births during the period April 2006 to December 2011 for all Japanese prefectures from the Ministry of Health, Labour, and Welfare (Ministry of Health, Labour, and Welfare of Japan, 2012). Japan keeps very accurate vital statistics with virtually complete coverage. The mandatory Japanese residential registration system (i.e., “toseki kohon”), moreover, allowed attribution of live births among evacuees to the mother's pre-Earthquake prefecture. We separated the birth records into those for the three most affected prefectures, the eight prefectures on the Island of Kyushu, and the remainder of Japan.
Tests
Our tests proceeded through the following five steps for each of the three test areas. First, we regressed the monthly number of male births on the number of female births to adjust the former for any phenomena that affect the monthly number of all births (e.g., the size of conception cohort, risk aversion and other characteristics of the reproductive population). Second, we used the method devised by Box and Jenkins (Box et al., 2008) to detect and model autocorrelation in the residuals of the model estimated in step 1. The method, Auto Regressive, Integrated, Moving Average (i.e., ARIMA) modeling, identifies which of a very large family of mathematical expressions best describes measurements made over time. The method assumes that the measurements passed through an unobserved filter, or model, that imposed autocorrelation (i.e., trends, cycles, and the tendency to remain elevated or depressed, or to oscillate, after high or low values) upon them. The approach looks for patterns in the data that narrow the search to likely models and then applies estimates of “fit” to identify the most likely candidate. The best fitting models estimate expected values that approximate what would occur if the circumstances that produced earlier observed values remain unchanged. The differences between these estimated values and the values observed measure the degree to which birth cohorts included fewer or more males than expected from the number of females and from autocorrelation.
Third, we constructed an indicator variable to specify the earthquake. We scored our earthquake variable 1 for March 2011 and 0 for all other months.
Fourth, we estimated the equation formed by adding the indicator variable constructed in step 3 to the model resulting from step 2. We specified the equation to estimate the association for the cohort born in the month of the Earthquake (i.e., March, 2011) as well as for cohorts born up to 9 months later (i.e., December, 2011). Selection in utero predicts a coefficient significantly less than 0 for cohorts born in June, July, or August, whereas reduced conception predicts such a coefficient in November or December. We note that the theories do not exclude each other, so significant coefficients could appear in both time “windows.” Both selection in utero and reduced conception of males imply a single-tailed test of significance for coefficients on the indicator variable because both predict negatively signed associations. Given that we would infer support for at least one of the arguments if any of five coefficients were significantly less than 0, we set significance conservatively at P < 0.025 to reduce the likelihood of a Type I error.
In the fifth step in our analyses, we tabled the expected and observed number of male births, as well as their differences, for March through December 2011 in each of the three test populations. The table identifies which differences the test equations estimated would occur by chance fewer than 2.5 times per hundred (one-tailed test).
RESULTS
Table 1 shows the means, standard deviations, and coefficients of variation for births in the three populations over the 69 test months (i.e., April 2006–December 2011). Relatively few births occur in the most affected and most distant areas compared to those in the remainder of Japan and monthly counts for the most affected prefectures yields a relatively low coefficients of variation. These circumstances suggest that estimates from the remainder of Japan will be the most stable and have the narrowest confidence intervals.
| Monthly Mean | Standard deviation | Coefficient of variation | |
|---|---|---|---|
| Most affected prefectures | |||
| Males | 1958.6812 | 130.8566 | 124.3348 |
| Females | 1860.4493 | 120.8679 | 127.8590 |
| Most distant prefectures | |||
| Males | 5752.6377 | 234.1287 | 204.0971 |
| Females | 5469.3188 | 222.5428 | 204.1476 |
| Remainder of Japan | |||
| Males | 38407.6957 | 1589.2388 | 200.7491 |
| Females | 36421.6812 | 1479.2452 | 204.5240 |
The first two steps of our analyses revealed that all six of the sex by area series exhibited strong seasonality. The three ARIMA models, shown in Table 2, that yielded the expected values of male births therefore included terms for integration, as well as a moving average parameters, at 12 months. Notation in these equations include the operator
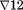 that indicates a series has been differenced (i.e., value at time t subtracted from value at t−12), as well as the backshift operator B12 or value of the variable it modifies at time t−12, and at or the difference between the expected and observed values of the series at time t.
that indicates a series has been differenced (i.e., value at time t subtracted from value at t−12), as well as the backshift operator B12 or value of the variable it modifies at time t−12, and at or the difference between the expected and observed values of the series at time t.
| Most affected prefectures |
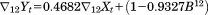 |
| Most distant prefectures |
 |
| Remainder of Japan |
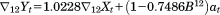 |
Table 3 shows the results of the final step in the analyses by listing the observed and expected values of the male births series, as well as their differences, for the 10 months March through December 2011. We estimated 30 differences for this table and would, therefore, expect 1 to be significantly less than 0 by chance (i.e., .025 × 30 = 0.75). We found 8. We found no differences significantly greater than 0. Consistent with a distance gradient, half of the significantly low observed values appeared in the most affected prefectures, one appeared in the most distant prefectures, and three in the large and populous area between.
| Birth month | Japan without most distant or most affected prefectures | Most distant prefectures | Most affected prefectures | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Observed males | Expected males | Difference | Observed males | Expected males | Difference | Observed males | Expected males | Difference | |
| March | 36,879 | 37,481.457 | −602.456a | 5,649 | 5,577.090 | 71.910 | 1,769 | 1,842.246 | −73.246 |
| April | 36,692 | 36,307.883 | 384.116 | 5,642 | 5,569.193 | 72.807 | 1,737 | 1,916.582 | −179.582a |
| May | 36,995 | 37,172.371 | −177.370 | 5,570 | 5,708.152 | −138.152 | 1,833 | 1,947.880 | −114.880 |
| June | 37,163 | 37,856.211 | −693.211a | 5,624 | 5,664.103 | −40.102 | 1,854 | 1,922.379 | −68.379 |
| July | 39,104 | 39,138.035 | −34.037 | 6,105 | 5,971.579 | 133.421 | 1,983 | 1,888.395 | 94.605 |
| August | 39,921 | 39,704.922 | 216.078 | 5,995 | 5,973.906 | 21.094 | 1,963 | 1,963.341 | −.341 |
| September | 39,556 | 39,525.953 | 30.045 | 5,993 | 6,040.566 | −47.566 | 1,910 | 1,955.135 | −45.135 |
| October | 37,729 | 38,620.645 | −891.644a | 5,677 | 5,867.469 | −190.469a | 1,822 | 1,906.578 | −84.578a |
| November | 36,021 | 36,167.020 | −146.021 | 5,624 | 5,534.012 | 89.988 | 1,612 | 1,695.844 | −83.844a |
| December | 36,125 | 36,525.594 | −400.593 | 5,837 | 5,898.641 | −61.641 | 1,545 | 1,645.261 | −100.261a |
- a P < 00.025; one−tailed test.
- Shown by distance from greatest destruction.
Results for the most affected prefectures appear consistent with the reduced conception hypothesis in that the birth cohorts plausibly conceived in March and early April, and born in November and December, included fewer than expected males. The timing of the six other birth cohorts with significantly fewer than expected males would, however, imply selection in utero because the earliest of these (i.e., those born in October in all three areas) would have been roughly in the second month of gestation at the time of the Earthquake. The remaining three cohorts with significantly fewer than expected males were born even sooner after the Earthquake implying that their male deficit resulted from selection in utero rather than from reduced conception of males.
We used the methods of Chang et al. (1988) to determine if outliers in our dependent variable could have inflated our confidence intervals and led us to underestimate the number of significant differences between observed and expected male births. No outliers were detected in any of the three models.
To help put the associations we estimated into context, we summed the “missing males” in the male-deficient Japanese birth cohorts during our test period. The eight significant deficits we discovered summed to 2,828 missing males. This represents about 2.2% of the 127,000 male births expected in the eight affected cohorts.
DISCUSSION
Our findings from the Great East Japan Earthquake of March, 2011 support the argument that population stressors can reduce male births via both reduced conception of males and selection against males in utero. We found support for reduced conception of males in that the population most affected by the Earthquake yielded fewer males 9 and 10 months after the disaster. We also infer support for selection in utero because six of the eight cohorts with missing males were conceived well before the Earthquake. These deficits appeared, moreover, among all three populations we studied.
We note, however, that the timing of male deficient birth cohorts differed from that offered in the literature arguing selection in utero. That argument implies that cohorts in the 16th–24th weeks of gestation should respond most to population stressors (Catalano et al., 2012b). The cohort born in June to the Japanese population without the most affected and most distant prefectures fell in that window. Although the relatively large population and relatively stable birth counts in this area may make this association compelling, it did not appear in the most affected or most distant populations.
The most consistent sensitive period, indeed across all three populations, suggested that gestations near the end of the first trimester lose males when subjected to population stressors. What we know of fetal loss suggests that much occurs in this period but presumably selects against the most profound of bars to fitness including chromosomal abnormalities and gross morphological abnormalities. The Great East Japan Earthquake data suggest, however, that an extreme population stressor at this point in gestation may reduce “tolerance” for abnormalities that would otherwise persist later into gestation and perhaps to birth.
We further note that only the population most distant and apparently least affected by the Earthquake did not show a male deficit within 2 months of the quake. Losses this late in gestation have traditionally been assumed to be too few to yield detectable responses to environmental stressors. Perhaps, however, the Great East Japan Earthquake so strongly stressed the population that late selection against small males increased significantly.
Our findings cannot arise spuriously from autocorrelation, including seasonality, in male births, or from phenomena that affected the number and outcome of gestations of females as well as males. We, moreover, believe that selection in utero explains the low sex ratios among cohorts in gestation at the time of the Earthquake more parsimoniously than does the rival explanation that the population choosing to conceive 1–7 months before the largest earthquake in Japanese history included more than otherwise expected persons with characteristics that make the conception of males less likely than that of females.
Our findings converge with those of Torche and Kleinhaus (2012) who studied the probability of male births in Chile following a strong earthquake in 2005. That study used validated vital statistics and rigorously controlled for autocorrelation as well as births in a comparison population. We, therefore, suggest that the most rigorous studies in the literature support the argument that selection in utero contributes to the unexpectedly few male births reported to follow earthquakes.
We note that events as destructive as the Great East Japan Earthquake occur only rarely. Our test, therefore, should be considered one of an extreme population stressor. Our findings do, however, converge with those (Catalano et al., 2012b) of similar reduction in male births after a much more common stressor (i.e., labor market contraction).
We further note that natural selection may have conserved mechanisms that eventually balance the effects of selection in utero (Bull, 1983). The effect of compensating mechanisms presumably appear when cohorts with highly biased sex ratios reach reproductive age (Carvalho et al., 1998). Tests to detect compensation for the effect of the Great East Japan Earthquake on sex ratios will have to wait at least two decades.
Further research might repeat our tests but separate the Japanese into populations based on theories of connectedness rather than on distance from epicenter. The research could separate the population into groups connected to the most affected prefectures not only by spatial proximity but also by telephone contacts, historic migration patterns, or transportation systems.
We acknowledge that an intuitively appealing complement to our test would have been to determine if registered (i.e., post 20th gestational week) spontaneous abortions and still births of males increased above expected levels in the affected prefectures after the Earthquake. Agencies responsible for registering such events have not, however, published monthly counts by prefecture. We do not, moreover, have access to the individual medical records from which we could estimate such counts. Even if we could have these records with identifiers removed, human subjects regulations concerning disclosure by knowing small area (i.e., prefecture) and brief time (i.e., month) would probably preclude us from using the data.
We leave to others to opine on whether a 2.2% reduction in male births below expected levels has short or longer-term implications for political, economic, or social phenomena. We speculate, however, that any statistically significant reductions due to selection in utero have clinical implications because they imply an increase in the population coping with the somatic and psychological sequelae of fetal death.
The implications of our findings for science seem clearer. Our results align with the claim that natural selection has conserved mechanisms that reduce the odds of a male live birth during stressful times by reducing the conception of males and by increasing the rate of spontaneous abortion among male fetuses. The conservation of mechanisms that can bias the sex of birth cohorts suggests that natural selection can quickly affect fundamental characteristics of contemporary populations.