

Strawberries under salt stress: ALA and ROS to the rescue
Abstract
Generating salt-tolerant plants that can cope with increasing soil salinity is a major goal of crop-breeding programs worldwide. Together with genetic approaches, research efforts are focusing on finding chemical modulators of salt tolerance. The exogenous application of 5-aminolevulinic acid (ALA) has been shown to improve salt tolerance in diverse crop species, but its mechanism of action is not properly understood. Wu et al. (2019) report that ALA treatment enhances reactive oxygen species (ROS) production in the roots of salt-stressed strawberry plants. Activation of several key ion transporters downstream to the ROS signal helps to sequester the toxic Na+ ions in the roots and protects the shoots against salt stress.
Sodium, which is an essential macronutrient for animals, is toxic to most land plants at millimolar concentrations. Why the majority of land plants evolved as salt-intolerant glycophytes is a much speculated topic. It is hypothesized that plants evolved to contain low sodium to make themselves less attractive to sodium-loving herbivores. Crop plants are especially vulnerable as selective breeding has diminished many salt tolerance traits (Cheeseman 2015). Salinity is certainly not a new threat to agriculture; it is believed that soil salinity was the major cause of the collapse of the agriculture-centric civilization of Mesopotamia nearly 4000 years ago. Nevertheless, recent increasingly erratic climatic conditions, including irregular rainfall and rising temperatures that can increase evapotranspiration, are certainly aggravating the rise of soil salinity. It is estimated that nearly 10 million hectares of irrigated lands are lost each year to rising salinity (Shahid, Zaman & Heng 2018). It is therefore not surprising that designing halotolerant crops is an area of active research, and this relies on the understanding of salt-induced toxic effects and mechanisms of salt management. The adverse effect of salt on plants is multifaceted. High concentrations of salt impose osmotic stress across the plasma membrane and also lower the water potential, thereby limiting water availability and subjecting the plants to drought stress. However, most of the toxic effects are exerted by the cytosolic presence of the Na+ ions, which compete with potassium as a cofactor of key enzymatic processes such as photosynthesis (Gupta and Huang 2014). At physiological levels, plants try to limit Na+ movement to the leaves to maintain photosynthesis and energy production. Upregulation of HKT1, a high-affinity potassium transporter, unloads Na+ from the xylem vessels and prevents its transport to the shoot. At the cellular level, the plants try to limit the concentration of Na+ in the cytoplasm by pumping it across the plasma membrane through the Na+/H+ antiporter SOS1 and by sequestering Na+ into the vacuole through another Na+/H+ antiporter, NHX1. How Na+ is initially sensed in the cell is still an enigma; however, a downstream calcium and ROS-dependent signaling pathway leads to upregulation of the aforementioned ion transporters (Yang and Guo 2018). In addition to maker-assisted breeding, efforts to improve salt tolerance have relied mainly on the overexpression of the ion transporters mentioned above. However, many recent studies have focused on the exogenous application of chemical compounds that can confer salt tolerance in a non-genetic way (Nguyen et al. 2018).
ALA is an important biochemical precursor in chlorophyll biosynthesis. Exogenous application, as well as genetic overexpression of ALA, has been shown to improve salt tolerance in a large number of crop species (Wu et al. 2018). However, the specific mechanism by which ALA conferred salt tolerance is not well understood. In this issue, Wu et al. (2019) show that ALA cotreatment promoted sequestration of Na+ in the roots and reduced its accumulation in the leaves of strawberry plants treated with 100 mM salt (compared to plants treated with only salt; Fig. 1A). The authors found that ALA cotreatment dramatically enhanced the salt-induced upregulation of SOS1, NHX1 and HKT1 in the root, which would account for the superior Na+ sequestration in the root (Fig. 1B). As salinity-induced ROS signaling plays an important role in the upregulation of these ion transporters, the authors explored whether
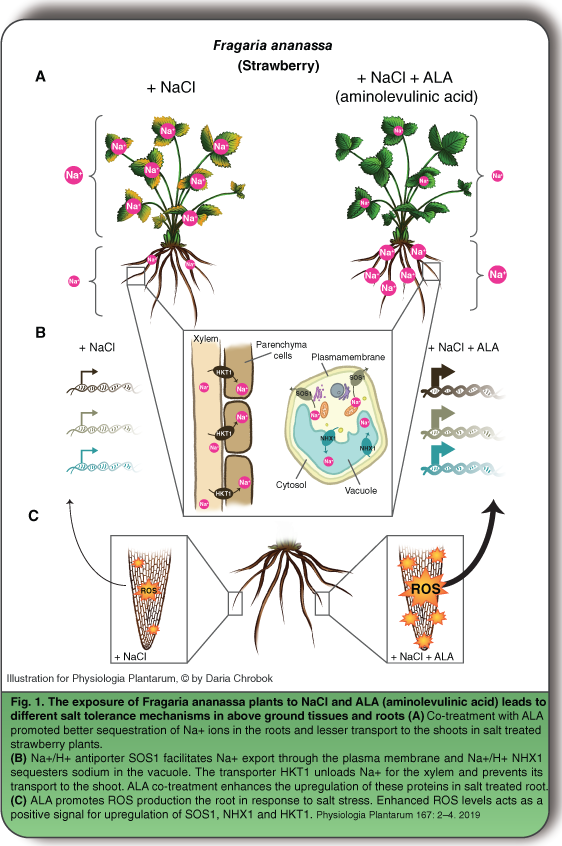
ALA treatment influenced ROS accumulation in the root and found that ALA treatment promoted salt-induced ROS accumulation (Fig. 1C). Intriguingly, the authors found that exogenously applied hydrogen peroxide had protective effects against salt stress similar to the ALA treatment. Moreover, treatment with diphenyleneiodonium, an inhibitor of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase-dependent ROS generation, abrogated the protective role of ALA against salt stress. These observations together highlight the importance of ROS generation and ROS signaling in mediating salt tolerance responses in plants.
The ion transporters (SOS1, NHX1, and HKT1) that act as key downstream effectors of salt tolerance have been known for nearly a decade. It has been convincingly demonstrated that constitutive overexpression of these proteins confers salt tolerance in plants (Yang et al. 2009). However, this approach has not gained enough impetus because of the transgenic nature of the resulting tolerant plants. Foliar application of chemicals provides an alternative and easier means to achieve salt tolerance in the field (Nguyen et al. 2018). Application of an endogenous plant metabolite such as ALA is promising for minimizing environmental concerns. However, in-depth studies are required to understand the mode of action of these compounds to assess their impact on overall plant health and productivity. Previous studies suggested a positive role for ALA in sustaining chlorophyll content under salt stress (Wu et al. 2018). The current study by Wu et al. (2019) will be an important step toward understanding and assessing the effect of ALA and will take us a step closer to commercial field applications of this compound.

