

Eucalypt seedlings are aided by phosphorus in the face of drought
Abstract
Drought is an increasingly common climatic event that can devastate ecosystems, as well as surrounding agricultural and forestry industries. Few places face this challenge more than Australia, where millennia of droughts linked to geography and climatic drivers, such as El Niño, have shaped the flora and fauna into forms predicated on resilience and economy. How an organism responds to these cyclic challenges is a combination of the inherent tolerance mechanisms encoded in their genome and outside influences, such as the effect of nutrients and symbiotic interactions. In this issue of Physiologia Plantarum, Tariq et al. (2019) describes how the presence of the element phosphorus can bolster the physiological and biochemical response of eucalypt seedlings to severe drought conditions.
The native range of Eucalyptus grandis extends along the eastern coastal and subcoastal regions that link New South Wales and Queensland. The timber of this tall, fast-growing eucalypt has an attractive pink hue (from which it gets its common name rose gum) and excellent wood properties. Consequently, E. grandis has received intense international interest and, today, is the most widely planted hardwood species worldwide and is used in a wide variety of applications, ranging from general construction and joinery to the production of pulp, charcoal and biofuel. Having such an expansive range of plantation exposes this economically valuable species to an increased incidence of abiotic stresses, particularly drought. Thus, recent efforts have been made to characterise the tolerance mechanisms used by E. grandis under drought conditions and investigate management practices that can be used to alleviate the worst of the effects.
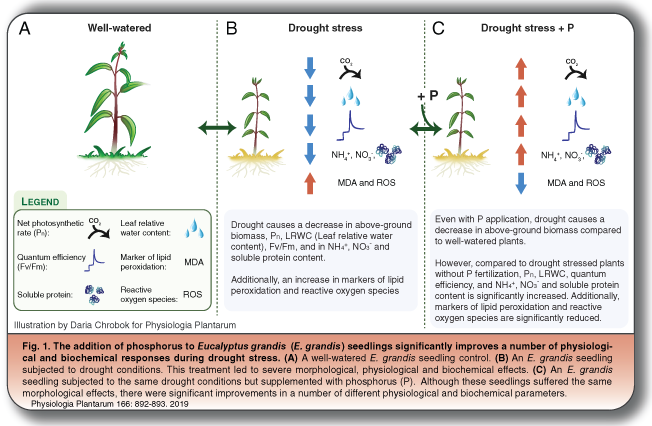
A number of recent studies have highlighted the role phosphorus (P) can play in mitigating drought stress in plants by promoting growth, especially in seedlings (Tng et al. 2014). This prompted Tariq et al. (2019) to investigate the morphological, physiological and biochemical response of E. grandis seedlings to two water regimes (simulating drought and non-drought conditions) when P was withheld or supplemented (Fig. 1). The researchers observed that, regardless of P application, drought conditions severely affected E. grandis seedlings, resulting in morphological effects in above-ground tissues, such as significant reductions in leaf and shoot biomass, leaf area and stem diameter, when compared with their well-watered counterparts. Root biomass was not affected by drought, which resulted in an overall decrease in shoot: root ratio, a situation far from ideal in forestry applications. These findings indicate that structural adaptations to drought stress in E. grandis, such as limiting water loss by reducing leaf area and maintaining root biomass to enhance water collection, cannot be relieved by P application (Fig. 1). However, this is only half the story as an examination of leaf physiology showed that P application in drought-stressed seedlings triggered significant improvements in net photosynthetic rates (Pn); leaf relative water content (LRWC – a helpful proxy for the water status of the plant); and the quantum efficiency of photosystem II, which provides a snapshot of the condition of the reaction centres of the photosynthetic apparatus in response to stress (Fig. 1C). Changes in the biochemistry of the seedlings' leaves were also catalogued, with P application in drought-stressed seedlings significantly improving NH4+, NO3− and soluble proteins content, which likely altered the leaf osmotic potential and contributed to the greatly improved LRWC observed. Finally, although drought treatment provoked a significant accumulation of reactive oxygen species and associated lipid peroxidation in seedlings, irrespective of P application, the magnitude of this stress response was significantly reduced when seedlings had access to P as evidenced by the observed reduction in lipid peroxidation products and O2− content. Despite the negative growth effects observed following the 90-day drought treatments, one could speculate that the improved phenotypic and biochemical parameters observed following P addition might eventually lead to long-term growth enhancements over the life of the tree.
Evidence from the fossil record and molecular phylogenetic analyses suggest eucalypts arose in what is now South America during the late Cretaceous or early Paleogene (Hill et al. 2016). During the late stages of the breaking up of the supercontinent Gondwana, their territory expanded and bridged into western Antarctica, New Zealand and southeastern Australia. Following this expansion, their range mysteriously shrank to such an extent that, by the 18th century, eucalypts were found only in Australia and a few adjacent regions of Indonesia and New Guinea. What precipitated the extinction of eucalypts outside of Australia is still a topic of debate; however, their obvious value to forestry industries the world over has seen rapid re-expansion, particularly into China, India, South Africa, Brazil and much of Mediterranean Europe. As this growth continues, studies such as Tariq et al. (2019) will be required to unravel the complex interplay between the tolerance potential of eucalypt species and the diverse environments they will come to inhabit. This could prove vital in informing future management practices to enhance their innate drought-tolerance adaptations with strategic nutrient additions to ensure the productivity and lasting future of this hardy tree.

