

The importance of type species and their correct identification: A key example from tintinnid ciliates (Alveolata, Ciliophora, Spirotricha)
Abstract
Types and the corresponding rules in the International Code of Zoological Nomenclature are crucial for taxonomy and are meant to provide nomenclatural stability. In the case of neotypification, especially diligent taxonomic work is required to retain continuity. In the present communication, we first outline the main principles of typification and neotypification. We then discuss a critical case, using a current example from the marine planktonic tintinnid genus Tintinnopsis Stein, 1867 (Alveolata, Ciliophora). This diverse and ubiquitous genus is nonmonophyletic, but its revision and the erection of new related genera is currently prevented by the uncertain affiliation of its type species.
RELIABLE taxonomy imposes a fundamental responsibility on all who are involved in taxonomic acts: the authors, the reviewers, and the journal editors. As a scientific community, we should maximize our efforts to prevent the publication of insufficient descriptions and redescriptions. In the first part of this paper, we provide a short summary of the type and neotype concepts contained within the framework of the International Code of Zoological Nomenclature (ICZN, 1999) and their application to protists. In the second part, we exemplify a case in the tintinnid genus Tintinnopsis Stein, 1867 (Alveolata, Ciliophora) and discuss problems and overall consequences for tintinnid taxonomy related to the proper identification of its type species, T. beroidea.
TYPES AND NEOTYPES
Types are crucial in taxonomy. For a given taxon, each of its nested ranks is anchored to a type (e.g., a specimen, species, or genus) that bears its defining features. The type is the link between the established scientific name and any future work on a given taxon. The nomenclature of metazoans and heterotrophic protists is governed by the International Code of Zoological Nomenclature (ICZN, 1999). The code includes several articles treating the “Principle of Typification.” One specimen (or a set of specimens) is the type of a nominal species group (subspecies or species; Article 61.1.2.), and one nominal species represents the type of a nominal genus group (subgenus or genus; Article 67.1.1.). Once a type is fixed, it is expected to be stable and to provide objective continuity in the application of the name (Article 61.1.3.) and the objective standard of reference (Article 61.1.). In brief, the type displays features that distinguish its nominal taxon from related ones (Article 61.1.1.). However, the Code does not require that a type displays all the diagnostic characters or that it averages in character states. Thus, type specimens sometimes lie at one extreme of the taxon boundaries and end resulting less “typical.”
Every taxon name meeting the criteria of Articles 10–20 of the Code (ICZN, 1999) must be considered in taxonomic work. This is markedly different from other biological disciplines, where researchers can judge and decide if publications are scientifically sound or should be considered insufficient.
Most species descriptions of heterotrophic protists were published in the 18th, the 19th, and the first half of the 20th century and contain, at maximum, those features accessible by the methods available at that time. By applying additional and more sophisticated techniques, the descriptions are continuously complemented, that is, the species are redescribed. This is a critical process, as taxonomists must carefully verify conspecificity between their specimens and the originally described ones. Frequently, and especially in the case of heterotrophic protists, this is a difficult task owing to the scarcity of characteristics provided by the original descriptions and the lack of deposited type specimens. Finding the balance between redescribing a species despite some uncertainties or establishing a new species (i.e., considering the existing species group name a nomen dubium) requires responsibility in order to prevent inflation of available names.
Redescriptions frequently involve neotypifications. Regarding protist taxonomy, perhaps, the most common reason for designating a neotype is an urgent need to define the taxon objectively and no name-bearing type specimen is believed to be extant (Article 75.1.; ICZN, 1999). The material should come “as nearly as practicable” from the original type locality (the geographical place of capture, collection, or observation) of the original name-bearing type (Articles 75.3.6., 76.). Since protists have been assumed to be much more widely distributed than metazoans, or even to be cosmopolitan, redescriptions including neotypifications are frequently based on material collected at sites distant from the type locality. According to some taxonomists, the wording “as nearly as practicable” provides the required degree of flexibility (Corliss, 2003; Curds et al., 2005; Song, 2003), while others suggest that protists should be completely exempt from the type locality requirement (Al-Rasheid, 2003; Dragesco, 2003; Foissner, 2002; Foissner et al., 2002). However, literature surveys and molecular studies have increasingly shown examples of biogeographic patterns in protists (Bass et al., 2007; Casteleyn et al., 2010; De Luca et al., 2019; Dolan & Pierce, 2013; Ganser et al., 2021; Richter et al., 2020; Santoferrara et al., 2018; Sommeria-Klein et al., 2020). Thus, neotypifications should only be performed based on specimens coming as nearly as practicable from the original type locality (or exceptionally be based on an authoritative redescription; see example below); otherwise, only voucher material should be deposited along with redescriptions (Curds et al., 2005). A neotypification should be accompanied by sound arguments demonstrating the compliance of Art.75.3.6. (ICZN, 1999). Previous neotypifications that violate this article should be accepted for the sake of taxonomic and nomenclatural stability, unless compelling new evidence exists.
The ICZN (1999) lists further conditions for qualifying a neotypification (Article 75.3.), for example, that the corresponding publication (i) clearly points out the exceptional need of clarifying any taxonomic problems and (ii) provides evidence that the neotype is consistent with what is known of the former name-bearing type from the original description. Furthermore, the Code recommends consultation with additional experts in the group under scrutiny prior to the designation of a neotype to prevent serious objection and taxonomic confusion (Recommendation 75B). Publication in an appropriate journal (i.e., one focused on protists, including their taxonomy) fosters additional involvement of experts (reviewers and editors) in critical taxonomic acts, such as descriptions, redescriptions, and neotypifications.
AN ELUSIVE TYPE SPECIES IN TINTINNID CILIATES
Ciliates and, in particular, tintinnids are among the most studied groups of protists. The loricae (shells or houses) of tintinnids are rather robust and thus easily collected by plankton nets. Accordingly, the about 1000 tintinnid species are characterized, almost exclusively, by lorica features, while only a comparatively small fraction has more recently been studied with regard to cytological characters. Until recently, the inability to produce permanent slides prevented the deposition of physical type material.
The present paper builds on previous recommendations for taxonomic work provided by a broad community of ciliate experts beyond the principles of the ICZN (1999). Focusing particularly on tintinnids (Santoferrara et al., 2016) or on ciliates as a whole (Warren et al., 2017), the following needs were emphasized: (i) accurate identifications by a detailed comparison with original descriptions and (ii) data on the relevant diagnostic features, that is, cell characters and DNA sequences (at least the 18S rRNA gene) in descriptions and redescriptions. For tintinnids, however, the lorica is still the only (or at least the most important) taxonomic character included in most original descriptions or authoritative redescriptions. Besides its shape, the diameter of the lorica opening (or the narrowest portion of the collar) is a crucial diagnostic character (Laval-Peuto & Brownlee, 1986), while the lorica length can be highly variable.
The marine planktonic tintinnid genus Tintinnopsis and its type species, Tintinnopsis beroidea Stein, 1867, meet a pressing need for clarification. Currently, the genus comprises about 140 species characterized by loricae having agglutinated foreign particles and a usually closed posterior end, but lacking a nuchal groove or an inner nuchal shelf (Kofoid & Campbell, 1929).
The genus is, however, nonmonophyletic in genetic and cladistic analyses (Agatha & Strüder-Kypke, 2014; Bachy et al., 2012; Santoferrara et al., 2017; Strüder-Kypke & Lynn, 2008) and includes species with phenotypic plasticity and/or crypticity (Santoferrara et al., 2013, 2015). Tintinnopsis-like species show at least five distinct patterns of somatic ciliature (Agatha & Strüder-Kypke, 2007, 2013, 2014; Gruber et al., 2018), and their 18S rRNA gene sequences are interspersed in at least twelve lineages in phylogenetic trees (Santoferrara & McManus, 2021). Furthermore, Tintinnopsis-like species frequently group together with tintinnids possessing hyaline (glass-like) loricae, suggesting that the agglutinated lorica represents either a plesiomorphic or a homoplasious character. Therefore, the genus urgently requires a revision and splitting into several distinct genera. To potentially give these clades a home in the future, Agatha and Strüder-Kypke (2013) recommended against synonymizing the Tintinnopsis-like genera Stylicauda Balech, 1951 and Rhizodomus Strelkow and Wirketis, 1950; further useful subgenera names were compiled by Aescht (2001). However, a revision of Tintinnopsis and related genera is currently impossible because (i) most of their species still lack cytological and molecular data, and (ii) it is still unclear which clade represents the “true” genus Tintinnopsis, that is, which one includes the type species.
Tintinnopsis beroidea was first described from the Baltic Sea by Stein (1867) without measurements or illustrations; a deposition of type material is unknown. The lorica is described as thimble-shaped and parabolically pointed posteriorly and consists of a hard, parchment-like, hyaline matrix with agglutinated mineral particles. The author also provided some cell features, using specimens that had supposedly abandoned their loricae. However, the diameter of the peristome (smaller than the body), polar kineties, shallow longitudinal furrows, and a semicircular macronucleus cast doubt on the identification of the aloricate cells as a tintinnid. Thus, only the information about the lorica features is reliable, but not detailed enough to allow an unequivocal identification (Jörgensen, 1912, 1924). The first detailed and illustrated redescription is from Entz (1884), using material from the Mediterranean Sea. Although again no deposition of type material is known, we agree with Jörgensen (1912, 1924) and Kofoid and Campbell (1929) in regarding this first redescription as authoritative. Entz (1884) wrote in German: “Hülse conisch-glockenförmig, mit weiter Mündung und spitz zulaufendem hinteren Ende; Länge und Breite schwankt je nach Individuen ziemlich bedeutend (figs. 1–3), die Länge zwischen 0,06–0,08, die Breite zwischen 0,05–0,06 mm” (lorica conical, campanulate, with wide opening and pointed posterior end; length and width vary considerably, namely, 0.06–0.08 mm in length and 0.05–0.06 mm in width). He considered the species the most common tintinnid in the Gulf of Naples at the Italian Mediterranean coast.
Despite its general utility, the most frequently used monograph for tintinnid identification, even nowadays, introduced confusion regarding T. beroidea (Kofoid & Campbell, 1929). The authors did not display any of the line drawings accompanying Entz's description (Entz, 1884), but, instead, an illustration published by Jörgensen (1924). Since Kofoid and Campbell (1929) did not mention the lorica size, its dimensions have to be inferred from this illustration and the magnification given. According to these data, the width of the lorica is calculated to be only 35 µm, which does not overlap with the 50–60 µm-range mentioned by Entz (1884). Hence, any identification based on the monograph of Kofoid and Campbell (1929) is misleading in the most reliable lorica character, that is, the opening diameter.
Tintinnopsis beroidea has been reported from almost all over the world (pers. database of SA), but this is generally based on doubtful identifications that reflect the confusion introduced by Kofoid and Campbell (1929) and other compilations (Zhang et al., 2012). Furthermore, the cell features and DNA sequences of T. beroidea are still unknown. Cytological data from specimens collected in Korean coastal waters and assigned to T. beroidea actually correspond to a different species with a lorica opening diameter of about 30 µm (Choi, 2015). A partial 18S rRNA gene sequence is labeled as T. beroidea in GenBank (EF123709), but its identification cannot be confirmed, as no accompanying publication exists. Moreover, a recently proposed redescription of T. beroidea (Bai et al., 2020) presents several taxonomic problems (detailed below) that prevent us from considering it authoritative.
The specimens studied by Bai et al. (2020) were collected from surface coastal waters off Qingdao, China, and thus from a different Longhurst ecological province than the Mediterranean Sea material (Longhurst, 2007). Tintinnids show clear biogeographical patterns at the genus level (e.g., Dolan & Pierce, 2013; Santoferrara et al., 2018), and possibly species-level differentiation among closely related Chinese and European variants (Ganser et al., 2021). A neotypification of T. beroidea based on specimens from China was not proposed, and it would not be justified based on the recommendation by Curds et al. (2005).
Further, we argue that the proposed redescription of T. beroidea (Bai et al., 2020) is based on misidentified specimens. The specimens from China have smaller opening diameters and their size range does not even overlap with that from the Mediterranean specimens (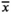 = 37 µm, 29–44 µm vs. 50–60 µm). Also, the lorica shape was misinterpreted, as Entz (1884) did not describe “the lorica as having a cylindrical collar and an obconical bowl with a pointed posterior end” (Bai et al., 2020). Although the lorica is the only link between the authoritative redescription (Entz, 1884) and extant populations, and despite the importance of the species as type of the genus Tintinnopsis, the information on lorica morphology provided by Bai et al. (2020) (“lorica campanulate, about 53–75 µm long, collar slightly flared with irregular rim with an opening 29–44 µm across. Bowl conical, diameter similar to opening, posterior end blunt, sometimes pointed”) is incomplete regarding the angles of the collar and the posterior end, the presence or absence of a spiraled or ring-shaped wall structure, and the percentage of specimens with a blunt or a pointed posterior end. Further, four stamp-sized micrographs and one line drawing display the lorica and its variability.
= 37 µm, 29–44 µm vs. 50–60 µm). Also, the lorica shape was misinterpreted, as Entz (1884) did not describe “the lorica as having a cylindrical collar and an obconical bowl with a pointed posterior end” (Bai et al., 2020). Although the lorica is the only link between the authoritative redescription (Entz, 1884) and extant populations, and despite the importance of the species as type of the genus Tintinnopsis, the information on lorica morphology provided by Bai et al. (2020) (“lorica campanulate, about 53–75 µm long, collar slightly flared with irregular rim with an opening 29–44 µm across. Bowl conical, diameter similar to opening, posterior end blunt, sometimes pointed”) is incomplete regarding the angles of the collar and the posterior end, the presence or absence of a spiraled or ring-shaped wall structure, and the percentage of specimens with a blunt or a pointed posterior end. Further, four stamp-sized micrographs and one line drawing display the lorica and its variability.
We also disagree with the synonymizations proposed by Bai et al. (2020). The authors synonymized Tintinnopsis acuminata Daday, 1887 with T. beroidea. Daday (1887) described his more slender variation acuminata of T. beroidea with a lorica length of 72–78 µm and an opening diameter of 45–50 µm. Although this range is continuous with the one of T. beroidea (50–60 µm), the difference in the opening diameter casts doubt on the conspecificity of both species and also with the specimens reported by Bai et al. (2020). Therefore, we agree with Kofoid and Campbell (1929), who raised the acuminata variant to species rank. Without any discussion, the authors also synonymized Tintinnopsis beroidea var. angustior Jörgensen, 1924 with T. beroidea. To avoid confusion, we favor a more conservative approach and the use of multiple lines of evidence for synonymizations. These taxonomic acts should be based on specimens matching the respective original descriptions, which have historically been based on the lorica. Given the known ambiguities of this structure, however, putative synonyms should also agree in cytological and molecular data. We also note that a hasty synonymization of species whose lorica dimensions (particularly the opening diameter) given by the original descriptions are only continuous or partially overlapping can eventually cause distinct unjustified deviations from the original species circumscriptions.
Finally, in their phylogenetic analyses, Bai et al. (2020) re-labeled as “T. beroidea” a sequence (JN831840) actually identified as T. acuminata (Santoferrara et al., 2013, 2017). Even if the sequences are identical, the usage of an accession number in combination with a taxon name different from that in GenBank is confusing, particularly, when it is based on an unjustified synonymization. To avoid future confusion, sequences MW011753 and MW011755 (Bai et al., 2020) should be re-labeled as Tintinnopsis sp. in GenBank until an identification can be achieved.
Based on the present evidence, the diagnosis of T. beroidea still exclusively comprises the lorica features described by Entz (1884). By all means, a revision of the genus Tintinnopsis Stein, 1867 and the establishment of new genera must be warded off at the current state of knowledge, as it would only cause serious harm to the stability of tintinnid taxonomy, systematics, and nomenclature. Such a taxonomic act, however, already took place (urn:lsid:zoobank.org:pub:3543CEC0-2490-416B-9120-9656645FE3A4) while the current Note was under review.
The collection of a neotype from the brackish waters of the Baltic Sea is prevented by the fact that neither an illustration nor measurements are available for the identification of the species established by Stein (1867). Actually, this was a case of nomen dubium until the redescription by Entz (1884). The decision of Kofoid and Campbell (1929) to give authority to Entz’s redescription (including the first illustrations and measurements) from the Mediterranean Sea is questionable based on the current regulations of the ICZN (1999). However, the outstanding monograph of Kofoid and Campbell (1929) has largely been used for tintinnid identification worldwide and represents a landmark for the taxonomy of these ciliates. Hence, striving stability in the application of the name T. beroidea necessitates sampling of Mediterranean specimens, ideally from the coastal waters of Naples, Italy, followed by an integrative approach, combining live observations, protargol staining, scanning electron microscopy, and sequencing of at least the 18S rRNA gene for redescribing the type species T. beroidea and a subsequent deposition of neotype material. Future tintinnid redescriptions and descriptions should provide many micrographs demonstrating the distinguishing features and the intraspecific variability and should consider all characters suggested by Santoferrara et al. (2016). Following the recommendations of the ICZN (1999; 72F) and Warren et al. (2017), type slides, neotype slides, and even voucher slides should be deposited in curated collections of major recognized museums to facilitate worldwide access.
CONCLUSION
To sum up, taxonomic acts such as descriptions and redescriptions should carefully consider the relevant diagnostic characters, the taxonomic literature, and the biogeography of the taxa of interest. The currently available data prevent a far-reaching revision of the nonmonophyletic genus Tintinnopsis. This is also partially true for the nonmonophyletic oligotrichid genus Strombidium and its type species S. sulcatum Claparède and Lachmann, 1859 originally described from Norwegian coastal waters and authoritatively redescribed from the coast of Brittany, France (Granda & Montagnes, 2003). At the current state of knowledge, far-reaching revisions of both genera would not aid the fundamental role of the ICZN in promoting nomenclatural stability.
ACKNOWLEDGMENTS
The study was financially supported by the Austrian Science Fund (FWF; I3268) given to SA. LFS is supported by the U.S. National Science Foundation (OCE-1924527) and the University of Connecticut.

