

Plasticity effect of rider–horse interaction on genetic evaluations for Show Jumping discipline in sport horses
Summary
To obtain a sport horse that excels in the highest levels of competition, breeders must take into account certain genetic and environmental factors that could influence the sport horse's performance, such as the rider–horse interaction (RHI). The main aim of this study was to describe this interaction in a genetic model by modelling it in relation to the horse's age. A total of 31,129 sport results from Spanish Sport Horses were used from a total of 1,101 animals evaluated, and these were grouped in three age levels and had been ridden by 606 different riders. Only riders who had ridden more than one horse (and vice-versa) were considered for the analyses. Five linear models with different random effects were analysed according to the covariates, the homogeneity/heterogeneity of the RHI and the relevant residual random effects. The model of best fit was then selected for the genetic evaluation of the animal. In general, models including the RHI effect (M2, M4 and M5) fitted better than the other models, and the best fit was obtained for M4 (with heterogeneous residual variance). The genetic variance increased constantly with age, whereas heritability showed a response on three intervals. This study revealed the varied evolution of the RHI with age, showing the different “plastic abilities” of this relationship.
1 INTRODUCTION
The main aim of any sport horse breed is to excel in competitions, whatever discipline they take part in (Koenen, Aldridge, & Philipsson, 2004). For this reason, breeders should take into account certain genetic and environmental factors that could influence the sport horse's performance (Bartolomé, Cervantes, Gómez, Molina, & Valera, 2008) not only during and between sports competitions (e.g., temperament, rider, location, weather), but also in horse breeding (e.g., sex, breed, stud conditions). Among the environmental factors that might influence sport performance, the rider is one of the most relevant, as they compete together in partnership with the horse. Hence, authors like Schöllhorn, Peham, Licka, and Scheidl (2006) reported that a well-balanced rider–horse interaction (RHI) resulted in better animal behaviour, reducing the stress associated with the competition and the possible accidents that it may cause. Unfortunately, although the importance of RHI is recognized in both sport competitions and other leisure activities, there are very few scientific papers that analyse its influence (Kearsley, Woolliams, Coffey, & Brotherstone, 2008). This could be due to one of the difficulties associated with the study of RHI, namely the structure and distribution of the data, as each rider usually rides either one horse or a very limited number of horses and each horse is usually ridden by only one rider during each exercise. These characteristics may produce a biased estimation of the RHI, which, together with the major disequilibrium included in the evaluation models, has excluded this factor from most genetic models of evaluation (Albertsdóttir, Eriksson, Näsholm, Strandberg, & Árnason, 2007) or, if included, it appears as a completely independent factor from the horse (Jaitner & Reinhardt, 1993). However, if this interaction is not taken into consideration, it could produce a bias in the genetic estimations that could further influence the response to selection, as highlighted in a previous study (Bartolomé, Menéndez-Buxadera, Valera, Cervantes, & Molina, 2013).
Among the Olympic disciplines performed by horses, Show Jumping is the one which measures the animal's sport performance most objectively (Valera et al., 2012), because it does not include subjective traits measured by a judge. However, several studies have demonstrated that the sport performance of young animals participating in this discipline is heavily influenced by other environmental factors, such as the rider or the age of participation (Bartolomé et al., 2008; Kearsley et al., 2008). In fact, to obtain the highest ranking in this discipline, a good, sound, trusting relationship between the horse and the rider is essential, as the influence of the rider on the horse's paces and jumps is critical (Munsters, Visser, van den Broek, & van Oldruitenborgh-Oosterbaan, 2012), as is the influence of the horse's reactions when faced with challenging situations (Visser et al., 2008).
Furthermore, changes in the horse's sport performance due to its interaction with the rider could be also considered as changes in the phenotype expression that depends on the environment, a biological property which has already been referred to as Phenotypic Plasticity (de Jong & Bijma, 2002; Via et al., 1995). These changes could be measured and quantified by two approaches: as a difference in the mean of the horse's phenotype (e.g., sport performance) for each environment considered (e.g., on each RHI), or the regression of a continuous environmental factor (e.g., considering each animal's age). While the former could be referred to as a Character State approach, the latter could be termed a Polynomial or Reaction Norm approach (Via et al., 1995). Furthermore, differences observed in either the rider's or the horse's sport ability with age and experience could lead to differences in the horse's sport performance. These changes of horse and/or rider could therefore be quantified with either of the two approaches previously explained (Character State or Reaction Norm), or with the plasticity effect of the RHI itself. Hence, the main aim of this study was to describe this interaction in a genetic model to estimate the sport horse genetic performance for Show Jumping competitions, modelling the relationship between the rider and the horse according to the horse's age, using norma-reaction techniques.
2 MATERIAL AND METHODS
2.1 Animals
In this study, Show Jumping competition data generated from the participations of the Spanish Sport Horse (CDE) breed were used. A total of 159 different competitions, held between January 2011 and May 2014, generated a total of 58,192 original participations from CDE horses. For the purposes of this study, only riders riding more than one horse and horses ridden by more than one rider were considered for the analyses. Hence, for this study, a total of 31,129 sport results were retained for analysis, involving 1,101 participating animals (479 males, 365 females and 257 geldings), aged from 4 to 16 years old. They were grouped in three age levels: 4 to 6 years old (19.5%), 7 to 10 years old (45.0%) and 11 to 16 years old (35.5%) and were ridden by 606 different riders. About 82.2% of the participating animals had at least five repeated records within a certain age level and 55.5% of them had participated in more than one age level. The connectedness of the database was tested from previous analyses using the methodology described by Thorén Hellsten, Jorjani, and Philipsson (2008).
The performance records for the Show Jumping discipline were measured through variable Positive Points (PP), which are a positive transformation of the penalty points obtained by the animal during the competition: the higher the penalties, the lower the PP. Jumping penalties were incurred for refusals and knockdowns. The values ranged from 0 to 200, with 200 representing the highest punctuation in the competition (0 penalties) and zero the lowest (maximum number of penalties).
The pedigree was completed with all the available information up to 4,253 animals, including 545 fathers and 725 mothers. Despite the fact that all the animals belonged to the Spanish Sport Horse breed, due to the mixed origin of this breed (Bartolomé, Cervantes, Valera, & Gutiérrez, 2011), they were grouped in five breed groups according to their country of origin: Germany; France; the Netherlands; Spanish Sport Horse; and Others (Bartolomé et al., 2011). Table S1 shows a descriptive analysis of the animals included in this study, according to gender and age.
2.2 Genetic models
Different linear models with different random effects were analysed to test the response of the dependent variable analysed (PP) to different environmental factors. All the environmental effects considered for the genetic models had been statistically significant (p < .05) in a previous GLM analysis (results not shown). The heterogeneous effects were analysed according to the three different age groups previously established. Next, statistical analyses were carried out for five different genetic models, which all shared similar fixed effects and a heterogeneous animal effect according to age. The genetic models were grouped according to the random effects included individual permanent environment (IPE), rider, RHI, Concat and Residual; as well as by the homogeneity/heterogeneity of two of them (RHI and Residual) according to age.
The random effects included were as follows: for Genetic model 1 (M1): individual IPE, rider, Concat and a homogeneous residual factor; for Genetic model 2 (M2): rider, Concat and homogeneous residual and RHI factors; for Genetic model 3 (M3): IPE, rider, Concat and heterogeneous residual factor; for Genetic model 4 (M4): rider, Concat, homogeneous RHI and heterogeneous residual factor; finally, for genetic model 5 (M5): Concat, heterogeneous residual and RHI factors.
 (M1)
(M1) (M2)
(M2) (M3)
(M3) (M4)
(M4) (M5)
(M5)where Y is the dependent variable PP. All the genetic models studied included three different fixed effects (fixedi): date of the event (159 levels); sex (three levels), grouped in males, females, geldings and breed (five levels), grouped according to the origin of the breed of the horse in Germany (Hannoverian, Holsteiner, Oldenburger, Westphalian), France (Anglo-Arab, Selle Français), the Netherlands (KWPN, Belgian Warmblood), CDE (Spanish Sport Horse) and Others, which included other minority sport horse breeds.
To represent the population response curve, the genetic models included a second order (r = 2) fixed covariate (b1) for the age effect. Փr is the Legendre polynomial coefficient; aj is a random genetic effects vector due to jth animal with data and its ancestors without records (4,253 registries in total), which is assumed heterogeneous, according to age groups, modelled with a Legendre polynomial of order r = 1 for M1 and of order r = 2 for M2; Hk is the random general environmental effect due to a kth Concat (2,413 levels), which was an effect designed according to the complexity of the Show Jumping events studied as the interaction of the following effects: “event” (competition day and place where it is performed), “category of the test” (difficulty level of the tests performed within the event), “scale of score” (type of scale used for awarding points in the test performed), “height of the fences” (maximum permitted height of fences in the test performed) and “type of test” (particular rules of the test performed) for a particular Show Jumping event; QI is a random effect of the Ith IPE due to the different participations of each animal (1,101 animals); Pm is a random effect of the mth rider (606 riders); In is a random effect produced by the nth RHI combination (2,903 levels), which was considered homogeneous in M2 and M4 and heterogeneous in M5, according to age groups; and eijkl is the random residual error, considered homogeneous in M1 and M2 and heterogeneous in M3 to M5, according to age groups. Z1, Z2, Z3, Z4 and Z5 are incident matrices with one or zero elements that connect the registries with the effects indicated in the subscripts.

where I represents identity matrices of one or zero orders which connected the yth records with the effects indicated in the subscripts. Variances were due to the animal ( ), the Concat effect (
), the Concat effect (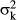 ), IPE (
), IPE (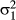 ), the rider (
), the rider (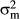 ), the horse–rider interaction (
), the horse–rider interaction (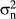 ) and the residual effect (
) and the residual effect ( ), respectively. A is the relationship matrix. In M1 and M2, a matrix of random regression coefficients of order one was included (Ka), which included an intercept with
), respectively. A is the relationship matrix. In M1 and M2, a matrix of random regression coefficients of order one was included (Ka), which included an intercept with 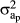 variance and a slope with
variance and a slope with 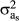 variance, representing both the general and specific genetic ability of the animal according to changes in age;
variance, representing both the general and specific genetic ability of the animal according to changes in age;  was the covariance between these two components.
was the covariance between these two components.
2.3 Statistical analyses
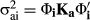 ; and covariances between ages I and j as
; and covariances between ages I and j as 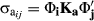 . The heritability for every ith age (
. The heritability for every ith age (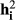 ) and genetic correlations (
) and genetic correlations (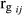 ) between ages i and j were obtained using the following classic formulas:
) between ages i and j were obtained using the following classic formulas:

Variance components were estimated for the five models considered. The variance of the heterogeneous animal effect was calculated as the variance of the intercept (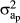 ), the variance of the slope (
), the variance of the slope (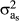 ), and the covariance between both (
), and the covariance between both ( ). In addition, the ratio between both variances (
). In addition, the ratio between both variances ( ) was also calculated. In the case of M3, M4 and M5, the variance of the heterogeneous residual effect was calculated for each age group: 4–6 years old (AG1), 7–10 years old (AG2) and 11–16 years old (AG3). For M5, however, both variances of the RHI and residual effects were heterogeneous and they were therefore calculated according to each age group analysed (AG1, AG2 or AG3).
) was also calculated. In the case of M3, M4 and M5, the variance of the heterogeneous residual effect was calculated for each age group: 4–6 years old (AG1), 7–10 years old (AG2) and 11–16 years old (AG3). For M5, however, both variances of the RHI and residual effects were heterogeneous and they were therefore calculated according to each age group analysed (AG1, AG2 or AG3).

where fb is the deviation of the fixed breed effect, aj = [apas] is a vector of the genetic function for each animal, estimated as the solution to every genetic model and  is the Legendre polynomial coefficient.
is the Legendre polynomial coefficient.
From the random regression matrix obtained for the genetic effect, the variance component of the intercept (Vint), and of the slope of the regression vector (Vslp), the covariance between both (Vi x slp) and the relation between both variances (Vslp/Vint) were calculated according to Kolmodin and Bijma (2004) to highlight differences in sport ability estimated across ages, according to the genetic model used.
To check their different responses, the evolution of the 150 best EBV animals was analysed for animals between 6 and 14 years old (those belonging to youngest and oldest age groups previously considered, respectively) and estimated for M3 and M4 (300 animals in total per genetic model). Finally, to check for different plastic responses of the RHI, the evolution of three animals with different behaviours across age for the RHI variance was estimated for M5 (with heterogeneity of the residual variance and the RHI variance due to age).
3 RESULTS
3.1 Genetic parameters and comparisons between genetic models
The genetic parameters obtained for the five genetic models analysed are presented in Tables 1 and 2. Variance components (Table 1) showed an increasing tendency by age in the animal effect except for M4 and M5, with percentages increasing from 5.5% to 7.9% for model 1; from 6.8% to 9% for M2; from 6.0% to 6.1% for M3 and decreasing from 9.1% to 7.9% for M4 and from 11.5% to 9.1% for M5. On the other hand, as regards homogeneous effects (IPE, rider and Concat), the latter accounted for the highest percentage of the variance, ranging from 16.5% for M2 to 23.3% for M5. In addition, the RHI factor explained a higher percentage of variance as a heterogeneous effect (M5) than as a homogeneous one (M2 and M4), showing a decreasing tendency with age in M5 (from 7.4% at AG1 to 3.1% at AG3). The residual effect was the factor which accounted for most of the variance for all the models, with values ranging from 51.2% at AG1 for model 5 to 70.9 at AG3 for M3. It also showed an increasing tendency with age in M3 to M5.
| Animal | IPE | Rider | RHI | Concat | Res | |
|---|---|---|---|---|---|---|
| M1 |
Vint = 114.5 Vslp = 34.03 Covi x slp = 61.28 Vslp/Vint = 0.297 |
54.10 | 199.26 | – | 453.12 | 1,835.52 |
| M2 |
Vint = 151.7 Vslp = 32.72 Covi x slp = 60.83 Vslp/Vint = 0.216 |
– | 186.32 | 67.85 | 451.81 | 1,820.89 |
| M3 |
Vint = 76.50 Vslp = 40.51 Covi x slp = 38.41 Vslp/Vint = 0.53 |
79.34 | 209.52 | – | 455.38 | AG1 = 1,103.85 |
| AG2 = 1,679.69 | ||||||
| AG3 = 2,293.53 | ||||||
| M4 |
Vint = 142.81 Vslp = 38.27 Covi x slp = 35.07 Vslp/Vint = 0.27 |
– | 194.17 | 81.77 | 453.54 | AG1 = 1,087.48 |
| AG2 = 1,661.19 | ||||||
| AG3 = 2,277.72 | ||||||
| M5 |
Vint = 217 Vslp = 28.3 Covi x slp = 51.2 Vslp/Vint = 0.13 |
– | – | AG1 = 157.60 | 636.01 | AG1 = 1,090.19 |
| AG2 = 158.40 | AG2 = 1,651.14 | |||||
| AG3 = 103.10 | AG3 = 2,290.64 |
- IPE, individual permanent environment; RHI, rider–horse interaction; Res, residual effect; Vint, variance of the intercept; Vslp, variance of the slope; Covi x slp, covariance between the intercept and the slope; Vslp/Vint, relation between the variance of the slope and the variance of the intercept; AG1 = 4–6 years old; AG2 = 7–10 years old; AG3 = 11–16 years old.
| Genetic models | p | LogL | AIC | BIC | h2 ± SEMa | R g |
|---|---|---|---|---|---|---|
| M1 | 7 | −4,098.53 | 8,211.06 | 8,269.44 | 0.067 ± 0.003 | Rg = 0.055–0.079 |
| M2 | 7 | −4,088.24 | 8,190.48 | 8,248.86 | 0.079 ± 0.003 | Rg = 0.068–0.090 |
| M3 | 9 | −3,753.46 | 7,524.92 | 7,599.98 | 0.061 ± 0.001 | Rg = 0.060–0.061 |
| M4 | 9 | −3,739.89 | 7,497.78 | 7,572.84 | 0.085 ± 0.002 | Rg = 0.079–0.091 |
| M5 | 10 | −3,809.63 | 7,639.26 | 7,574.33 | 0.047 ± 0.004 | Rg = 0.036–0.078 |
- a For all genetic models, the mean heritability was shown, as the genetic value was heterogeneous due to age.
According to the genetic parameters shown in Table 2, informative criteria such as LogL, AIC and BIC were calculated to compare the genetic models, despite the fact that these criteria give only general guidelines for interpretation and do not serve as a formal analysis for the best fit. In general, models including the RHI effect (M2, M4 and M5) fit better than the other models, with the best fit obtained for M4 (with heterogeneous residual variance). As regards the heritability values, those models including the RHI effect as homogeneous showed higher heritability values than those without or showing a heterogeneous RHI effect, with 0.085 for M4 compared to M5 (h2 = 0.047). The relative importance of the rider and the RHI over the phenotypic variance were calculated for the genetic model of the best fit (M4) (Figure 1). This figure showed a decreasing tendency due to age for both rider and RHI effects, with a stronger effect for the rider, decreasing from over 8% to below 5% of the total variance, than for the RHI, which decreased from 3.5% to 2% of the total variance.

The evolution of the RHI variance and heritability across age was represented in Figure 2, as well as for the genetic model of best fit (M4). The genetic variance increased constantly with age, while heritability showed a response in three intervals: the first interval between 4 and 7 years old, where no changes were registered except for a slight decreasing tendency; a second interval, from 7 to 11 years old, showing the lowest heritability values for all the ages; and finally, the third interval, from 11 to 16 years old, which shows an increase until it reaches the maximum response of 0.07 at 16 years old.

3.2 Study of the plastic response
Figure 3 shows the genetic models of best fit (M3 and M4) comparing models with and without homogeneous RHI (M4 and M3, respectively), to check the plastic ability of the animals’ breeding values evaluated at different ages. The results showed that the Pearson's correlations found between and within models (r values) were high and positive, ranging from 78.3%, between the EBV calculated with M4 at AG1 and the EBV calculated with M3 at AG3, to 80%, between the EBV calculated with M3 at AG1 and the EBV calculated with M4 at AG3. These values still leave a percentage of over 20% for differences in the animals’ genetic response animals and thus, making possible the response for genetic improvement.

Finally, Figure 4 shows the heterogeneous RHI and residual variances for M5 to check the plastic ability of the RHI with age. This figure shows, for three different animals randomly chosen from the database, the average genetic potential at different ages (estimated as the sum of the horse's genetic value plus the effect of the interaction of a particular horse with a given rider) and the range for which that potential is established, according to the different riders with whom he has competed. Animal 5807 showed an increasing tendency with age with EBV values ranging from 5 to almost 8 units over the mean, with a high range according to the rider–horse combination for every age level. Animal 12758 showed an overlap with animal 5807's results for ages 4 to 6, with a decreasing tendency with age, ranging from 4.5 to 3.5 and 4 EBV. On the other hand, animal 12690 showed lower EBV values, which ranged from 3 to 2.5 and 2.8 at more advanced ages, showing almost no variability of the RHI for every age level.

4 DISCUSSION
Several studies, including a previous study of young Spanish Sport Horses (Bartolomé et al., 2013), have shown the need to take the rider effect into account (either as rider or RHI) in genetic models (Peeters, Closson, Beckers, & Vandenheede, 2013), however, it has been rarely considered in genetic analysis (Kearsley et al., 2008), mainly due to the difficulty involved in estimation, where a considerable imbalance occurs in the analysis models. Therefore, to obtain more robust and consistent conclusions, the data structure in this study included only riders who had ridden more than one horse and horses ridden by more than one rider. Furthermore, following to a previous study (Bartolomé et al., 2013), only multiple trait animal models were compared, to focus on the RHI effect.
According to the results obtained in our study, when considering the variance components, genetic models including the rider and the RHI factors accounted for a higher percentage of the variance than the additive genetic effect itself, hence corroborating the importance of taking into account these factors in the genetic models, as highlighted on a previous study (Bartolomé et al., 2013). However, in this study, the evolution of the RHI variance, the rider genetic variance and the heritability were studied over an age trajectory, hence focusing on a different plastic effect that had not been analysed before.
The results found in the present study showed an evolution in the animals’ EBV with age according to the genetic model used. It therefore highlighted the animals’ “plastic ability” according to age, finding either robust or plastic animals for Show Jumping competitions, according to the RHI influence.
This evolution of the animals’ EBV with age has been also reported previously by other authors (Thorén Hellsten et al., 2006; Viklund, Braam, Näsholm, Strandberg, & Philipsson, 2010) , who reported high correlations between parameters measured in young animals and later in life, hence highlighting different performances due to age.
The heritability values found in our study were similar to those from Bartolomé et al. (2013), although in this study, the RHI and rider effects were not studied over an age trajectory, ranging from 0.069 to 0.343 for the heterogeneous animal models.
According to the comparison criteria used for the fit (AIC, BIC and LogL), M4 (heterogeneous residual variance due to age including RHI effect) showed the best fit. This result supports previous conclusions reported by other authors about including the rider effect in the genetic models (Bartolomé et al., 2013). This corroborates the importance of including the rider effect in genetic evaluations in horses (either as the rider itself or as the RHI) to give unbiased estimations of breeding values.
When accounting for the plastic ability of the rider and the RHI effects (Figure 1), as well as the genetic variance and the heritability (Figure 2) over age, a decreasing trend was shown by both effects which contrasted with an increasing trend shown by genetic variance and heritability. This highlights an underlying learning and experience acquired by the animal with age so that the older the animal becomes, the less influenced it is by the rider in its sport results. On the other hand, it becomes more accustomed to the rider, and rider–horse fit constitutes a lower environmental influence, which, in turn, increases the heritability estimation. According to McGreevy and McLean (2010), a horse's response to the rider's requirements will depend on the rider's cues and the horse's prior learning of an appropriate response to these cues. Hence, the longer the horse has been learning the cues, the more appropriate its response to the rider. Furthermore, the exposure to novel stimuli or challenging situations was also perceived as less stressful by horses with a good horse–rider “match” (Munsters et al., 2012), which could be achieved easily with age as most horses were ridden by the same group of riders during their lives.
Figure 3 showed the plastic ability of the animals. Firstly, it can be seen that, regardless of the genetic model used (with or without RHI interaction), a considerable coefficient of variation of the EBV was shown at either a more advanced age (when selecting the best for younger animals) or at a younger age (when selecting the best for older animals). According to de Jong and Bijma (2002), phenotypic plasticity is the property of the organisms of a certain genotype to develop systematically different phenotypes in different environments. Hence, genotypes showing highly variable phenotypes across environments are considered “plastic,” whereas genotypes showing little variability across environments are considered “robust.”
Reaction-norm models (where the phenotype is described as a continuous function of an environmental variable) have been used previously in different animal species, as cattle (Mota et al., 2016), pigs (Verardo et al., 2017) or even wild animals (Sae-Lim, Komen, Kause, & Mulder, 2014). In horses, this methodology has been used for different purposes, such as to study the influence of heat stress on performance (Gómez, Menéndez-Buxadera, Valera, & Molina, 2015) or sport performance in Show Jumping competitions (Bartolomé et al., 2013). Gómez, Menendez-Buxadera, Valera, and Molina (2010) also described different sport performances in Trotter horses due to race distances, finding that the estimates of breeding values showed different evolutions for each animal according to race distance, thus differentiating between sprinters and stayer horses.
When CDE horses were analysed, we found animals with a plastic response where the horse's sport performance ability changed, either increasing or decreasing with age, so that the older the animal became, the more significant its interaction with its environment. On the other hand, there were other horses that showed a robust response, as their sport performance remained more or less the same through age, implying that the environment had very little influence on their sport performance. Thus, our results imply the possibility of selecting for different types of phenotypic response according to the interest of the breeders: selecting for robustness when the aim is to maintain an average performance no matter what environment conditions they participate in, hence maintaining similar performance level in Show Jumping, regardless of age and external circumstances; or for plasticity, obtaining the best possible results for certain environmental conditions even at the expense of presenting unequal results in other circumstances, different results according to the environmental influence, hence reaching their best performance at a certain age and depending on external circumstances. Despite the fact that the first option seems more desirable for these kind of animals, which have to face such a wide range of environments in each competition, animals with a good plastic response are usually able to achieve higher results given the right environment (Gómez et al., 2010), thus being more desirable as candidates to selection. It has to be considered that in general, riders tend to ride the same horse/group of horses throughout their sport career, hence it would be better selecting for a plastic group of horses that could excel with a particular rider/group of riders which are going to ride them. Furthermore, as competitions for adult animals award the highest prizes, it could prove more beneficial to select for plastic “late” horses, which could excel at the end of their sporting lives, instead of plastic “precocious” horses that only excel when they are young.
When the distribution of the breeding values in Figure 3 was analysed, a higher number of animals aged either 6 or 14 years old obtained breeding values of over 10 points when using M4 than with M3. These results also highlight the importance of choosing the right genetic model to compute the breeding values.
On the other hand, Figure 4 showed the plastic ability of the RHI with age, illustrating the evolution of RHI with age for three randomly chosen animals. It highlighted a clearly different evolution of performance potential of the CDE horse at Show Jumping competitions with age, when ridden by a certain group of riders. The EBV ranges within each age level indicate the variability of the horse's response to different riders at a certain age. The narrower the EBV ranges for each age level, the less influence of the rider on the horse performance with age and hence, the more robust the horse's response.
To illustrate this, Animal 5807 showed a genetic potential that increased with age, but with a high variability of this potential according to the rider–horse combination for every age level. It showed a moderate plastic response per age, and a different fit with riders throughout its sporting life. When compared with animal 12758, an overlap was shown in the younger age groups, indicating that a genetic selection at this earlier stage would make no difference in these horses as, no matter what rider rides these horses, they all show a very similar Show Jumping performance. However, when they grow up, their sport performance changes according to the rider chosen (hence increasing the rider's influence on the genetic potential of the horse), with a clearly higher genetic potential for animal 5807 than for animal 12758. On the other hand, animal 12690 showed a more stable, robust response as it not only changed its EBV with age, but also showed no variability in the RHI at each age level. This horse therefore showed no differences in its EBV due to the rider and hence, a very low RHI. This means that no matter the rider, the horse would probably show a very stable sport performance—perhaps not as high as the other horses showed, but stable over time—which could represent quite an interesting characteristic to select for. On the other hand, it should be noted that these robust results could also be due to homogeneous riding levels between the riders riding those horses.
According to Visser et al. (2008), the rider's personality does affect the cooperation with more emotionally reactive horses. These horses may therefore show a plastic response with age, and according to their interaction with the rider, could have a more reactive temperament than those showing a robust response with age. On the other hand, Peeters et al. (2013) found that compliant and partly-compliant animals (i.e., horses that matched different riders) showed lower stress levels than non-compliant horses (i.e., horses that did not match different riders). Taking into account the fact that stress can influence a horse's sport performance by either improving or worsening their sport results (Bartolomé & Cockram, 2016), horses showing a plastic response in our study may also be non-compliant horses, hence showing more stress in their interactions with different riders, which could bias their Show Jumping results. On the other hand, robust animals were more compliant, thus maintaining a more constant performance in their interactions with different riders. On the other hand, it has to be taken into account that, in practice, most genetic evaluations in horses are biased by the fact that best riders tend to select the best horses to ride at competitions. Thus, this horse–rider match would be already influenced by rider's preselection of a suitable horse (Wolframm & Meulenbroek, 2012). However, our approach highlights the evolution of this relationship with the age of the animal.
These results highlight the importance of choosing a suitable horse–rider match and selecting for the most appropriate type of interaction (plastic or robust) could be key in estimating the sport horse's performance correctly.
In conclusion, our results reveal not only the importance of including the RHI as a factor in the routine genetic models for Show Jumping genetic evaluations, but also how this influence changes with the animal's age (and hence experience), thus biasing the animal's results during its sporting life if this fact is not considered. It also influences the rider and RHI variances, diminishing the influence of the rider on the breeding values as the animal's age (and therefore experience) increases.
However, the results obtained in this study should be taken with caution in most equine populations, as they are directly applicable in those populations where the number of horses ridden by a single rider and the number of riders ridding a single horse is low in relation to the total participating population. Otherwise, the estimation of the RHI effect might be biased.
On the other hand, our results also imply the possibility of selecting for different types of phenotypic responses according to the breeders’ interest, selecting either for robustness or for plasticity for Show Jumping performance and of taking into account the interaction of the horse with the rider according to age. Furthermore, it highlights the importance of choosing the right horse–rider match, as our results showed either a plastic or robust behaviour of a horse's performance ability depending on the rider. Thus, selecting for the most suitable ability of this interaction (plastic or robust) could be key for correctly estimating a sport horse's performance.
ACKNOWLEDGEMENTS
The authors wish to thank the Department of Resource Management and Animal Feeding (Subdirección General de Conservación de Recursos y Alimentación Animal) of the Spanish Ministry of Agriculture, Food and Environment for their support. We would also like to thank the Spanish Sport Horse Breeders’ Association for their collaboration.

