

Movement patterns of Antillean manatees in Chetumal Bay (Mexico) and coastal Belize: A challenge for regional conservation
Abstract
Information from 15 satellite-tracked Antillean manatees (Trichechus manatus manatus) was analyzed in order to assess individual movements, home ranges, and high-use areas for conservation decisions. Manatees were captured in Chetumal Bay, Mexico, and tagged with Argos-monitored satellite transmitters. Location of the manatees and physical characteristics were assessed to describe habitat properties. Most manatees traveled to freshwater sources. The Maximum Area Size (MAS) for each manatee was determined using the observation-area method. Additional kernel densities of 95% home range and 50% Center of Activity (COA) were also calculated, with manatees having 1–3 COAs. Manatees exhibited two different movement patterns: remaining in Chetumal Bay, and long-distance (up to 240 km in 89 d). The residence time in Chetumal Bay was higher for females (89.6% of time) than for males (72.0%), but the daily travel rate (0.4–0.5 km/d) was similar for both sexes. Most of the COAs fell within Natural Protected Areas (NPA). However, manatees also travel for long distances into unprotected areas, where they face uncontrolled boat traffic, fishing activities, and habitat loss. Conservation of movement corridors may promote long-distance movements and facilitate genetic exchange.
The highest abundance of Antillean manatees (Trichechus manatus manatus) in the Mexican Caribbean is at the southern part of the Yucatan Peninsula, particularly in Chetumal Bay (Morales-Vela and Olivera-Gómez 1997). Chetumal Bay was designated as a Natural Protected Area (NPA) for manatees by Mexico in 1996 (Gobierno del Estado de Quintana Roo 1996) and as a Wildlife Sanctuary by Belize in 1998 (National Park System 1998). However, manatees in Chetumal Bay, as elsewhere, continue to face threats that include habitat degradation and loss, watercraft collisions, incidental catch/accidental take in fishing gear, pollution, human disturbance, natural disasters, and potential illegal hunting (Morales-Vela and Padilla-Saldívar 2009a). Therefore, identification of manatee movement patterns and high use locations may improve management of these protected areas. Although gross patterns of habitat use and movements of manatees have been inferred for different seasons in Chetumal Bay from aerial surveys (Olivera-Gómez and Mellink 2005), findings from radio tracking can provide considerably more detailed knowledge about seasonal ranges, movement patterns and pathways, and seasonal ecology of manatees (e.g., Deutsch et al. 2003, Arraut et al. 2009).
We conducted a manatee telemetry study in Chetumal Bay, Mexico, with the objectives of determining: (1) individual movement routes and home ranges, (2) high-use areas of individual manatees and their characteristics, and (3) site fidelity and residency patterns of manatees in Chetumal Bay.
Materials and Methods
Study Area
This study took place in Chetumal Bay (17º52'–18º50'N, 87º50'–88º25'W), a 2,560 km2 brackish shallow lagoon that has several fresh water inflows, including the Hondo River and New River. This coastal lagoon has a mainly flat, shallow bottom (3.2 m on average) marked with many small sinkholes called “pozas” (Carrillo et al. 2009a). The dry season in the region extends from February to June. The rainy season is from July to October, usually with a mid-summer moderate drought in August. Precipitation then decreases from November to January, corresponding to a period with several cold fronts (Carrillo et al. 2009b).
Capture, Tagging, and Radio-tracking
Fifteen manatees (eight females and seven males) were captured and tagged in northwestern Chetumal Bay between October 2004 and May 2007 as part of a long-term research project addressing manatee health status, movements and demography (Morales-Vela and Padilla-Saldívar 2009b). The locations and number of manatees captured were: La Barra (n = 10), Guerrero Lagoon Channels (n = 3), Ría Crik (n = 1), and Caleta Calderas de Barlovento (n = 1) (Fig. 1). Manatees were captured and tagged using the techniques detailed in Morales-Vela and Padilla-Saldívar (2009b). Sex, length, weight, and other standard measurements were recorded. The tag assembly (Platform Terminal Transmitter [PTT], Telonics, Inc., Mesa, AZ) consisted of a buoyant, cylindrical housing containing an Argos (Service Argos, CLS, Saint-Agne, France) satellite transmitter linked with a GPS receiver (Telonics TMT 240 or TMT 460). The tag assembly included a flexible nylon rod attached to a belt that fit snugly around the manatee's peduncle (Reid et al. 1995). The TMT 240 units were programmed to acquire GPS fixes at 30 min intervals, when they stored latitude, longitude, time, date, fix quality, and fix status. TMT 460 units were programmed to acquire GPS fixes at 20 min intervals, and stored latitude, longitude, time, and date. Data were uplinked to Argos satellites for remote acquisition, and also stored in nonvolatile memory for download after tag recovery. The tags were retrieved either by recapture of manatees or by recovering devices that had detached.

Telemetry Data Analysis and Habitat Assessment
Telemetry data were edited to remove fixes acquired before and after each animal's tagging period, and to eliminate location outliers. TMT 460 units provided regular and extremely precise (<10 m) spatiotemporal location data (Sheppard et al. 2006). However, TMT 240 units were somewhat less precise and resulted in some outliers on land. These outliers were removed prior to analysis.
We used GPS locations to calculate Centers of Activity (COA) for each individual, defined as the geographical location of the area of greatest activity within the home range (Dixon and Chapman 1980). Preliminary probability kernel contours of 50% were created for each tagged manatee using telemetry data obtained from the beginning of the study until August 2007 and the Animal Movement Analyst Extension (AMAE) of ArcView 3.3 (Environmental Systems Research Institute, Inc., Redlands, CA) (Hooge and Eichenlaub 1997). Benthic parameters (depth and bottom type) were measured in nine areas within the preliminary COAs (Fig. 1). The size of each area and the number of benthic sampling points by area and season are presented in Table 1. The substrate was categorized in situ as soft or hard depending on the absence or presence of rocks. We measured water temperature and salinity data using a YSI 6600 multiprobe. Bathymetric data were provided by the Laboratory of Physical Oceanography of ECOSUR, Chetumal.
| Area | Name | Size (km2) | No. of sampling points/season | Total no. of points sampled |
|---|---|---|---|---|
| 1 | Rio Creek | 17.95 | 25 | 75 |
| 2 | Cayo Venado | 23.04 | 32 | 96 |
| 3 | La Barra | 2.53 | 4 | 12 |
| 4 | Guerrero Lagoon Channels | 1 | 3 | 9 |
| 5 | Guerrero Lagoon | 5.7 | 8 | 24 |
| 6 | Tamalcab | 66.81 | 92 | 276 |
| 7 | Centro | 14.62 | 20 | 60 |
| 8 | Calderitas | 21.93 | 30 | 90 |
| 9 | Calderas de Barlovento | 9.32 | 13 | 39 |
| Total | 162.9 | 225 | 675 |
Home range estimates can be biased due to differences in tracking times among individuals. Therefore we assessed the number of tracking days that was sufficient to calculate home range for this group of manatees. We established the day when each manatee reached the Maximum Area Size (MAS), assuming that it corresponded to the home range of the individual determined using the observation-area method (Lehner 1996). This analysis is based on the principle that as the number of observations increases, the rate of increase in the area visited decreases, so that an asymptotic curve results. We used only the longitude (Y) data as a proxy for range size because manatee movements mainly occurred along coastlines and Chetumal Bay's principal axis is north-south. For each tracking day, we estimated the maximum range size as the distance between the most distant locations along the N-S axis (based on daily mean Y coordinates) (Deutsch et al. 2003). Then, we plotted maximum range size vs. tracking day, and noted the point after which additional locations resulted in a minimal increase in range size (Harris et al. 1990). In the home range analysis we only considered those individuals in which the curve reached an asymptote. We used the fixed-kernel density method to generate definitive COAs and home ranges at the 50% and 95% contours, respectively. The kernel density method considers the distribution of locations within the home range and determines one or more core-activity areas, or COAs (Worton 1989). Manatee residence in Chetumal Bay was assessed based on the percentage of days that each individual was detected within the perimeter of the study area (Jessopp et al. 2004, Bruce et al. 2005).
Results
Fifteen manatees were tracked from 4 to 301 d (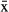 = 78 ± 78.1 d), yielding 32,957 fixes in total, for an average of 27.3 ± 16.2 fixes/d (Table 2). The fix success rate varied from 4.1 to 54 successful fixes/d. The mean failure rate (failed fixes/potential fixes * 100) was 61% (95% confidence interval [CI] = 60.7–61.4), and was higher for manatees instrumented with TMT 240 units (
= 78 ± 78.1 d), yielding 32,957 fixes in total, for an average of 27.3 ± 16.2 fixes/d (Table 2). The fix success rate varied from 4.1 to 54 successful fixes/d. The mean failure rate (failed fixes/potential fixes * 100) was 61% (95% confidence interval [CI] = 60.7–61.4), and was higher for manatees instrumented with TMT 240 units (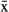 = 68.4%, 95% CI = 68.0–69.0) than for those with TMT 460 units (
= 68.4%, 95% CI = 68.0–69.0) than for those with TMT 460 units (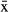 = 46.3%, 95% CI = 45.5–47.1).
= 46.3%, 95% CI = 45.5–47.1).
| ID | Sex | Capture site | Length (cm) | Weight (g) | GPS tag model | Beginning | Ending | Tracking time (d) | Potential fixes | Actual fixes | Fix success rate | % Failure |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 16 | F | La Barra | 283 | 365 | Gen III | 12/10/2004 | 27/12/2004 | 76 | 4,469 | 642 | 8.45 | 85.63 |
| 35 | M | La Barra | 267 | 335 | Gen IV | 26/03/2006 | 21/04/2006 | 26 | 1,872 | 1,418 | 54.54 | 24.25 |
| 36 | M | La Barra | 292 | 412 | Gen IV | 27/03/2006 | 22/01/2007 | 301 | 21,487 | 13,973 | 46.42 | 34.97 |
| 38 | M | La Barra | 228 | 215 | Gen III | 29/03/2006 | 28/05/2006 | 60 | 4,349 | 2,390 | 39.83 | 45.04 |
| 39 | M | La Barra | 280 | Unknown | Gen III | 18/04/2006 | 13/09/2006 | 148 | 7,094 | 1,541 | 10.41 | 78.28 |
| 40 | F | La Barra | 268 | 378 | Gen III | 11/10/2006 | 26/10/2006 | 15 | 2,626 | 571 | 38.07 | 78.26 |
| 42 | F | Caleta Calderas | 286 | 360 | Gen III | 13/10/2006 | 09/11/2006 | 27 | 2,762 | 911 | 33.74 | 67.02 |
| 44 | F | La Barra | 280 | Unknown | Gen III | 04/12/2006 | 30/01/2007 | 70 | 5,038 | 1,026 | 14.66 | 79.63 |
| 45 | F | La Barra | 268 | Unknown | Gen III | 19/12/2006 | 23/12/2007 | 4 | 195 | 46 | 11.00 | 77.44 |
| 47 | M | Guerrero Lagoon Channels | 304 | 371 | Gen III | 21/03/2007 | 27/04/2007 | 37 | 1,884 | 881 | 23.81 | 53.24 |
| 48 | M | Boca Ria Creek | 268 | 360 | Gen III | 22/03/2007 | 23/07/2007 | 123 | 8,565 | 2,486 | 20.21 | 70.97 |
| 51 | F | Guerrero Lagoon Channels | 293 | Unknown | Gen IV | 26/04/2007 | 05/05/2007 | 9 | 608 | 472 | 52.44 | 22.37 |
| 52 | F | La Barra | 312 | Unknown | Gen IV | 18/05/2007 | 04/10/2007 | 139 | 9,152 | 3,811 | 27.42 | 58.36 |
| 53 | F | La Barra | 285 | Unknown | Gen IV | 24/05/2007 | 20/06/2007 | 27 | 1,300 | 110 | 4.07 | 91.54 |
| 54 | M | Guerrero Lagoon Channels | 256 | Unknown | Gen III | 24/05/2007 | 09/09/2007 | 108 | 5,188 | 2,679 | 24.81 | 48.36 |
Manatees exhibited two different patterns of ranging behavior. In the first type (seven females and two males) manatees remained in Chetumal Bay during the entire tracking time (Fig. 2). In this group, two manatees (ID-35♂ and ID-51♀) exclusively used the freshwater system of Guerrero Lagoon, while the others moved throughout the bay. In the second type (one female and five males) manatees displayed long-distance movements out of Chetumal Bay (Fig. 3). Once outside of the bay, tagged manatees remained close to the coast, sometimes traveling up rivers. The longest linear travel registered was made by ID-52♀; she traveled 240 km (straight line) in 89 d from the capture point in La Barra, Chetumal Bay to south of Placentia Lagoon in Belize. Her trip was not continuous; she stopped at several areas en route. She stayed 1 mo in Belize before returning to Chetumal Bay, where her tag was removed after 139 d of tracking.


One male (ID-36♂) also traveled south twice from Mexico to Belize. This male was captured and tagged on 27 March 2006. He started to travel south 22 d after capture, reaching Southern Lagoon (Belize) in 10 d. He remained there about 2 mo, and on 4 July traveled north, reaching Chetumal Bay in 6 d. Three months later he returned to Southern Lagoon. The tag detached in January 2007 near the northwestern coast of Belize City.
Eleven manatees travelled up major freshwater sources, including Guerrero Lagoon (visited by nine manatees), Hondo River (five manatees), New River (two manatees), Northern River (one manatee), Belize River (one manatee), Northern Lagoon (two manatees), and Southern Lagoon (two manatees) (Fig. 2, 3).
The areas that were most consistently used by the 15 manatees in Mexico were Guerrero Lagoon, the center of Chetumal Bay, and near Chetumal City. In Belize high use areas were Northern Lagoon, Southern Lagoon and its southern coast, and the northwestern coast of Belize City (Fig. 4, 5).


Manatees captured in Chetumal Bay exhibited a diversity of ranging behaviors. Manatee residency within Chetumal Bay, as measured by the percentage of time spent there, was higher for females (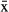 = 89.6%, 95% CI = 89.0–90.3) than for males (
= 89.6%, 95% CI = 89.0–90.3) than for males (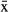 = 75.5%, 95% CI = 74.8–76.2), with only one female traveling out of the bay (Fig. 3). However, this pattern could have been influenced by tracking time, which was longer for males. The average speed was 2.74 km/h; the daily travel rate was similar for both sexes (
= 75.5%, 95% CI = 74.8–76.2), with only one female traveling out of the bay (Fig. 3). However, this pattern could have been influenced by tracking time, which was longer for males. The average speed was 2.74 km/h; the daily travel rate was similar for both sexes (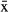 ± SD = 0.4 ± 0.13 km/d for females and 0.5 ± 0.12 km/d for males). Most of the tracked males traveled outside the coastal lagoon and most of the females remained in Chetumal Bay (Fig. 2, 3). There were two important exceptions to this rule. Male ID-39 did not leave the bay during the 148 d tracking time, and had a small home range. The length of this individual was 280 cm (adult-sized). Additionally, the female ID-52 ranged farthest from the bay and had the largest home range of all individuals. There were no evident differences in size, behavior, or health status that separated these two individuals from the others.
± SD = 0.4 ± 0.13 km/d for females and 0.5 ± 0.12 km/d for males). Most of the tracked males traveled outside the coastal lagoon and most of the females remained in Chetumal Bay (Fig. 2, 3). There were two important exceptions to this rule. Male ID-39 did not leave the bay during the 148 d tracking time, and had a small home range. The length of this individual was 280 cm (adult-sized). Additionally, the female ID-52 ranged farthest from the bay and had the largest home range of all individuals. There were no evident differences in size, behavior, or health status that separated these two individuals from the others.
Manatees reached their MAS in 29 ± 31 (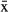 ± SD) d for females and 57.4 ± 36.2 d for males (Table 3). MAS was correlated with tracking time r = 0.69). We could not estimate the day of MAS for two males. On the basis of these results, we developed the home range analysis only with data from manatees that were tracked longer than 30 d (females) or 60 d (males). Eight manatees met this criterion; they had one to three COAs (Fig. 4, 5).
± SD) d for females and 57.4 ± 36.2 d for males (Table 3). MAS was correlated with tracking time r = 0.69). We could not estimate the day of MAS for two males. On the basis of these results, we developed the home range analysis only with data from manatees that were tracked longer than 30 d (females) or 60 d (males). Eight manatees met this criterion; they had one to three COAs (Fig. 4, 5).
| ID | Sex | Daily travel rate (km/d) | Day of maximum area size (MAS) | %Residency in Chetumal Bay | Home range size (km2) 95% kernel | COA size (km2) 50% kernel | Number of COAs |
|---|---|---|---|---|---|---|---|
| 16 | F | 0.24 | 72 | 100 | 202 | 26,14 | 2 |
| 35 | M | 0.65 | NE | 100 | – | – | – |
| 36 | M | 0.53 | 90 | 66.71 | 2,706.16 | 332.89 | 1 |
| 38 | M | 0.50 | NE | 95.69 | 1,185.93 | 195.04 | 3 |
| 39 | M | 0.65 | 103 | 100 | 288.23 | 42.95 | 1 |
| 40 | F | 0.46 | 14 | 100 | – | – | – |
| 42 | F | 0.35 | 40 | 100 | – | – | – |
| 44 | F | 0.21 | 8 | 100 | 267.79 | 18.16 | 1 |
| 45 | F | 0.50 | 3 | 100 | – | – | – |
| 47 | M | 0.37 | 26 | 54.48 | – | – | – |
| 48 | M | 0.35 | 36 | 63.27 | 3,351.61 | 676.3 | 1 |
| 51 | F | 0.50 | 10 | 100 | – | – | – |
| 52 | F | 0.47 | 79 | 17.08 | 5,156.73 | 680.82 | 2 |
| 53 | F | 0.58 | 8 | 100 | – | – | – |
| 54 | M | 0.53 | 32 | 24.07 | 2,637.71 | 320.56 | 2 |
The COAs were located in areas with depths of 1–4 m. In general, bottom type was mainly soft (83.5%) with few mixed (7.1%) or hard areas (9.4%). Hardness of the bottom was mainly due to the presence of rocks. The mean water temperature was 29.4 ± 1.7°C with lower values during November–January. Salinity ranged from 0.02 to 13 psu (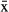 ± SD = 8.4 ± 2.7 psu), with a temporal pattern that corresponded to seasonality of rainfall.
± SD = 8.4 ± 2.7 psu), with a temporal pattern that corresponded to seasonality of rainfall.
Discussion
Ecological Characteristics of Areas of High Manatee Use in Chetumal Bay
Individual manatees typically occupied few, relatively small core areas. Travel routes followed the coastline, with activity centers at shallow (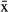 ± SD = 2.2 ± 1.1 m) estuarine habitats such as coastal lagoons or river mouths. Use of shallow water close to the coastal margins (Fig. 2, 3), agrees with previous studies on manatee depth preferences (Lefebvre et al. 2001), but their movements covered all the depth ranges found in Chetumal Bay (2–5 m). It is likely that manatees selected these coastal and shallow water areas more often because forage, such as submerged aquatic vegetation (Short and Neckles 1999, Havens 2003), or shoreline emergent vegetation (Castelblanco-Martínez et al. 2009), are generally found at such places.
± SD = 2.2 ± 1.1 m) estuarine habitats such as coastal lagoons or river mouths. Use of shallow water close to the coastal margins (Fig. 2, 3), agrees with previous studies on manatee depth preferences (Lefebvre et al. 2001), but their movements covered all the depth ranges found in Chetumal Bay (2–5 m). It is likely that manatees selected these coastal and shallow water areas more often because forage, such as submerged aquatic vegetation (Short and Neckles 1999, Havens 2003), or shoreline emergent vegetation (Castelblanco-Martínez et al. 2009), are generally found at such places.
There is evidence that in addition to the generally accepted habitat needs of manatees, such as foraging and access to freshwater, resting areas for cows and calves, and travel corridors between activity centers also may be important habitats (Packard and Wetterqvist 1986, Gannon et al. 2007). In Drowned Cayes, Belize, the presence of resting holes was an important explanatory variable for manatee sighting probability (LaCommare et al. 2008). The substrate in and around the resting holes tended to be a silty mixture of mud and sand, which was easily compressed and resuspended (Bacchus et al. 2009). In northern Chetumal Bay sand, soft silt and clay sediments are present (de Jesús-Navarrete et al. 2000, Molina-Ramírez 2007), indicating that Chetumal Bay also includes optimal habitat for bottom-resting activities.
Movements, Home Range, and Residency
Earlier studies of the movements of VHF tracked manatees in Chetumal Bay suggested that males traveled longer distances and showed less site fidelity than females (Morales-Vela and Medrano-González 1999). Our sample size (number of individuals and number of tracking days) was not large enough to show significant sexual differences in home range. Furthermore, the data obtained were strongly biased by differences in tracking time, which is a common issue in marine mammal telemetry research (Reid et al. 1995, Aarts et al. 2008). However, it is interesting that other authors noticed sexual differences in manatee ranging movements. In the St. Johns River, Florida, male manatees have been shown to travel particularly long circuits, presumably searching for receptive females (Bengtson 1981). Thus, males might maximize their own individual reproductive success by ranging longer distances than females. However in Chetumal Bay daily travel rates of male and female manatees were similar.
Studies carried out in Florida have shown that many manatees return to Crystal River each winter. The mean return rate has been calculated at around 90%, indicating strong site fidelity (Powell and Rathbun 1984). Other data from Florida show a greater site fidelity in females than in males (Rathbun et al. 1990). Manatees in this study were tracked for less than one year, so questions remain regarding annual movements and site fidelity at Chetumal Bay. However, the data obtained from ID-36 provided valuable multiseason information, showing seasonal movements similar to those described by Deutsch et al. (2003) for medium-distance migrants in Florida. The environmental driver of the migrations in the study area is unknown. Migrations of Florida manatees are closely tied to changes in water temperature (Deutsch et al. 2003), but annual water temperature in Chetumal Bay is from 24°C to 31°C (Morales-Vela et al. 1996) year-round, far above the temperature at which manatees experience cold stress, which has been estimated to be about 20°C (Bossart 2001, Bossart et al. 2003). Movement of Florida manatees are also likely to be influenced by seasonal fluctuation in standing crops (biomass), energy content, epiphyte load, and proximate constituents of their principal forage plants (Deutsch et al. 2003). In Florida productivity and growth of seagrass are positively related to irradiance and water temperature, but in tropical latitudes seasonal variation in productivity tends to be less pronounced (Lee et al. 2007).
Plasticity in Foraging Strategies
Manatees are generalists and extremely opportunistic herbivores, exploiting the most abundant forage species in their habitat, including macrophytic algae (Ledder 1986, Mignucci-Giannoni and Beck 1998, Borges et al. 2008). The macrophyte community of Chetumal Bay is composed of 47 species: the algae Batophora spp. is the dominant genus, followed by freshwater species such as the algae Chara spp., the herbaceous vascular plant Najas marina, and the brackish-water seagrass Ruppia maritima (Espinoza-Ávalos et al. 2009). The wide distribution of West Indian manatees, and the great diversity of habitats they occupy (Lefebvre et al. 2001), result in great variation in foraging strategies (Deutsch et al. 2003, Reich and Worthy 2006). This means that although the presence of abundant vegetation is a determining factor for their distribution and habitat use, the foraging strategy (e.g., diet composition, temporal and spatial variation.) might vary geographically based on local characteristics.
For this sample of 15 manatees, the residency periods (number of days) in high-use areas inside of Chetumal Bay were shorter than those of high-use areas in Belize. Spatial patterns in foraging may result from animals responding to patterns of forage availability, resulting in proportionally more time spent in nutrient-rich areas (Bailey et al. 1996), such as seagrass patches. Seagrass in Chetumal Bay is widely distributed but with low density and biomass (Espinoza-Ávalos et al. 2009). In contrast, vegetation is abundant in large beds throughout Southern Lagoon and dominated by the seagrasses Halodule wrightii and Ruppia maritima (Hamilton 2009), which are scarce in Chetumal Bay. In a nutrient-poor area such as Chetumal Bay, short residency times might be expected, and were often seen. Nonetheless, previous studies of VHF radio-tagged manatees showed that two adult females with calves stayed within small home ranges in Chetumal Bay for nearly three years (Morales-Vela and Medrano-González 1999).
Implications for Conservation
Our results provide details about movements of manatees in the Yucatan region of the Caribbean. Based on the similarity of the movement patterns to those of Florida manatees, it is likely that these patterns may occur throughout their range. Long-distance movements of manatees across their range are important because they might facilitate genetic exchange. Most manatee populations in the Caribbean are relatively isolated and/or reduced due mainly to anthropogenic threats such as illegal hunting, entanglement and habitat fragmentation (Self-Sullivan and Mignucci-Giannoni 2008). This situation increases the chances for reductions in genetic diversity. Lower genetic variation can lead to increased population sensitivity to infectious agents (O'Brien 1994). Reduction in genetic diversity also has an impact on individual fitness and flexibility in coping with environmental challenges. As a consequence, mean fitness of populations decreases and lowers their resilience and long term adaptability (Lacy 1997).
Chetumal Bay in Mexico is designated as a Manatee Sanctuary (Gobierno del Estado de Quintana Roo 1996) and habitat in Belize is protected as Corozal Bay Wildlife Sanctuary (National Park System 1998). Most of the manatee COAs fell within these protected areas. Only ID-52♀ and ID-54♂ had COAs located in nonprotected areas. Results from aerial surveys have also been consistent with these observations (Morales-Vela et al. 2000). However, the majority of individuals left the NPAs to travel up freshwater sources. Only two of these areas (Laguna Guerrero and Southern Lagoon), have a protected status. Although selected NPAs in the Caribbean have been recognized as outstanding refuges for manatees, some individuals may travel long distances venturing up into the river systems. Some of these unprotected areas (e.g., Hondo River, Northern River, and Belize River) are threatened by uncontrolled boat traffic, eutrophication, and pesticide pollution (DC-M and BM-V, pers. obs.). Conservation of corridors could promote long-distance movements and facilitate genetic exchange (Chetkiewicz et al. 2006). Creating a single large regional protected area with levels of protection varying within it according to manatee use may be worth considering (Reeves 2000).
Continuing long-term telemetry studies of Antillean manatees is important in order to assess their home ranges and temporal patterns of movement. This information is useful not only for the understanding of manatee ecology and behavior, but also as a conservation tool. Ecology and behavioral research play a vital role in efforts to understand how anthropogenic activities affect manatee populations and habitats. Interpreting what is normal or abnormal cannot be done without knowing the timing of major life-history events, energy requirements, movement or migration patterns and behavior (Williams 2011). For instance, studies of trans-boundary movements can help identify whether manatees are migrating or dispersing (Quintana-Rizzo and Reynolds 2010). So far, our data have confirmed long-distance movements of manatees but due to limitations in tracking time, failed to show a consistent seasonal pattern. Future tracking of Antillean manatees throughout annual cycles will better define their spatial and temporal patterns of movement. With hundreds of other species of concern in the Caribbean, protection of habitat for manatees as an umbrella species may also indirectly protect the larger ecological communities of the region.
Acknowledgments
Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government. This research was funded by SEMARNAT/CONACyT (Project 2002-C01-1128). CONACyT also gave a Ph.D. study grant to DNC-M. Dolphin Discovery Group and Sirenian International Inc. gave additional economic support. Temporal captures and radio-tagging of manatees in Mexico were done under permission of SEMARNAT-Dirección General de Vida Silvestre (permit documents NUM/SGPA/DGVS/03144, 04513, 03670/06 and 03675/07). Thanks to Rafael Estrada-Anaya, Gerardo Rodríguez-Martínez, Marco Benitez-García, Daniel Robelo, Manuel Robelo, Humberto Bahena-Basave, Mauro Sanvicente-López, and many volunteers for their collaboration in manatee captures and tracking. The authors thank field assistants Isaí Madriz-Villanueva, Cristian Barajas, Jose Olivares-Escobedo, Isabel Mercadillo-Elguero, Alejandro Aragón-Moreno, and José Angel Cohuo Collí for their help in ecological data collection. Two anonymous reviewers of U.S. Geological Survey improved the manuscript with their comments and advice. Thanks are due to two anonymous reviewers and to Dr. Tom O'Shea for their very valuable comments on the manuscript.

