

Impact of artificial photoperiodism on the colony development of the bumblebee Bombus terrestris
Abstract
This study investigated the effect of the photoperiodic regimes 0 h light : 24 h dark (LD 0:24), LD 8:16, LD 16:8 and LD 24:0 at 28°C and 50% Relative Humidity (RH) on the colony development of hibernated (2.0, 2.5, 3.0, 3.5 and 4.0 months) bumblebee queens. The queens which had hibernated for 3.0 months and which were reared in a LD 8:16 photoperiod showed the highest rate of colony initiation (88.2%), colony foundation (67.6%) and progeny queen production (38.2%). The photoperiod LD 8:16 also produced the shortest period of colony initiation and colony foundation. The highest number of sexual males (171.2 ± 12.2) and queens (91.2 ± 9.9) were produced in the colonies when 3.5 and 3.0 month hibernated queens were kept in an LD 8:16 photoperiod. The results show that light regime and hibernation duration affect colony characteristics of Bombus terrestris.
INTRODUCTION
The colony of Bombus terrestris L. is annual and founded by a single queen. At the end of summer, males and new queens develop. After mating, the young queens leave the nest and fly for several weeks in daylight before they burrow into the ground where they spend the winter in hibernation. In spring time, queens end their hibernation and are exposed to daylight when they seek out a suitable site to form a colony. Queens continue to forage in daylight when they feed their first batch of larvae and even when the first workers appear (Heinrich 1979; Tasei & Aupinel 1994). Despite this behavior, several authors have reported rearing bumblebee queens in the dark (Van Heemert et al. 1990; Asada & Ono 2000).
Photoperiodic cues change the life history of organisms (Danks 2002), and insect diapause is usually affected by photoperiod (Danilevskii 1965). Philogene and McNeil (1984) described the effects of photoperiod on insect development and reproduction. Tasei (1994) reported that artificial light (8 h light : 16 h dark (LD 8:16)) influenced the oviposition frequency and egg-laying periods of narcotized bumblebee queens. Artificial photoperiodism showed a stimulating effect on oviposition and sexual production of overwintered B. terrestris queens (Tasei & Aupinel 1994). In Adalia bipunctata L., egg-laying was initiated in a photoperiod LD 13:11 and inhibited by LD 12:12 (Iperti & Prudent 1986). Hodek (1974) stated that Aelia acuminate L. laid eggs earlier in an LD 18:6 photoperiod, and Myrmica rubra L. workers laid eggs more readily in an LD 16:8 photoperiod than an LD 8:16 or in dark conditions (Hand 1983). Yoon et al. (2003) suggested that artificially overwintered B. terrestris queens showed a higher rate of oviposition, colony foundation and production of sexuals in an LD 8:16 photoperiod compared to an LD 0:24 or an LD 16:8 photoperiod.
Diapause is an important state of bumblebee biology, and it is the main obstacle in mass rearing. Bumblebee queens experience diapause from 6 to 9 months, and termination of diapause is governed by spring temperature (Alford 1969). Other than temperature, the most common and reliable cue affecting the diapause of insects is the photoperiod (Leather et al. 1993). However, several studies have been described to rear colonies of B. terrestris in captivity providing artificial photoperiods, yet satisfactory colony development maintaining different photoperiods and hibernation durations is still speculative. Knowledge of the optimum photoperiod and hibernation duration will greatly enhance the laboratory rearing process of bumblebee. The present study was undertaken to evaluate the relationship effect of photoperiod and hibernation duration on the colony development of B. terrestris.
MATERIALS AND METHODS
Origin of queens and activation
Bombus terrestris L. queens hibernated in peat at 4°C and 80% moisture for a period of 2.0, 2.5, 3.0, 3.5 and 4.0 months were obtained from the mass rearing system of the Department of Agricultural Biology, Kyungpook National University, Daegu, Korea. For the purpose of activation, queens were introduced into flight cages (40 cm × 40 cm × 60 cm) having illumination facility, and provided with ad libitum frozen pollen grain of the Korean Kiwi, Actinidia arguta Planch, and sugar solutions (1.5:1, w/v). The cages were kept for 1 week in separate growth chambers at a standardized temperature of 28°C (Duchateau & Marien 1995), 50% RH and photoperiodic regimes of LD 0:24, LD 8:16, LD 16:8 and LD 24:0. Photoperiodic treatments were administered with 220/240 V fluorescent white light (Philips MX204HF136, 40 Hz, λ = 0.96–0.17) at an intensity of ∼700 lux on the floor of the flight cages. The intensity of light was measured with a digital lux meter and the photophase was regulated with timer.
Stimulation of oviposition and colony foundation
Activated queens which rubbed their abdomen on the ground of the flying cages were kept separately in small transparent plastic boxes (16 cm × 11 cm × 7 cm), and given frozen Korean Kiwi pollen grain in a 4 cm diameter Petri dish and sugar solution (1.5:1, w/v) in a perforated plastic tube (35.3 cm × 35.3 cm × 35.3 cm). The rearing boxes were kept in the respective photoperiodic chambers. After 2 days, 1–2 day-old frozen queen pupa and one anesthetized B. terrestris worker were added to each rearing box to stimulate the queens for oviposition (Duchateau & Marien 1995). Each pupa was horizontally fixed with paraffin on hard drawing paper, so that the pupa could not roll and the queen could sit for oviposition. Pollen grains were supplied daily into the rearing boxes and sugar tube was changed weekly. The frozen pupa and anesthetized worker were substituted once a week unless the queen laid eggs on the cocoon or on the hard drawing paper. When the queen laid eggs, the narcotized worker was removed from that rearing box. The boxes were kept in these conditions until the workers of the first brood cells emerged. For further development, the colonies were shifted to larger colony boxes (27 cm × 18 cm × 13 cm) that were connected to sugar tanks (30 cm × 20 cm × 4 cm) by a cotton filter. The colonies completed their development in a dark room at 28°C and 50% RH. In the colonies, bees were fed with frozen pollen bars prepared from 50% honey solution and fresh Korean Kiwi pollen powder, and unlimited amounts of sugar solutions (1.5:1, w/v).
Observations
From the day after first introduction of the queen pupa, the colonies were checked regularly. The following observations were noted under red light:
- 1
Colony initiation (number of days between the day after first pupa supply until the laying of first eggs).
- 2
Colony foundation (number of days between the day after first pupa supply until the emergence of first brood worker).
- 3
Number of worker produced from the first brood egg cells.
- 4
Total number of sexuals (males and queens) produced in the colonies.
On the basis of the size of brood, we kept approximately 100–200 workers in the nest boxes. To check competition between queen and workers and among workers, excess workers were removed after emergence of male or queen cocoons. Consequently, young males and queens were collected daily. Data of the total number of sexual males were recorded until 30–40 days after the competition point (worker oviposition, egg eaten or more than two open egg cells).
Statistical analysis
Data were analyzed by two-way anova. The mean values of colony initiation, colony foundation, number of first brood workers and progeny queen production data were separated by Duncan's multiple range test. Tukey HSD post hoc test was employed to separate the mean values of the data of total number of males and queens. All the analyses were performed using SPSS, version 12.1 (SPSS, Chicago, IL, USA).
RESULTS
Rate at which queens initiated and founded colonies
In all the treatments, queens laid eggs and the proportion of queens-initiated colonies varied from 61.8 to 88.2% (Fig. 1). Significantly the highest oviposition rate (88.2%) was found in 3.0 month hibernated queens reared in LD 8:16 conditions. Two-way anova of the data indicated that photoperiodic regimes showed significant effects on colony initiation (F3,660 = 2.6, P < 0.05), whereas hibernation duration, and the interaction effect of photoperiod and hibernation duration did not show significant effects (hibernation duration: F4,660 = 1.4, P = 0.23; interaction: F12,660 = 0.3, P = 0.99) on colony initiation.

Rate (%) of Bombus terrestris queens initiating colonies according to photoperiodic regimes and hibernation durations (n = 34 queens). Bars with no common letter are significantly different (P ≤ 0.05, Duncan's multiple range test). Photoperiods (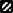 ) 0 h light : 24 h dark (LD 0:24), (
) 0 h light : 24 h dark (LD 0:24), (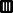 ) LD 8:16, (
) LD 8:16, (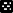 ) LD 16:8 and (
) LD 16:8 and (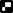 ) LD 24:0.
) LD 24:0.
Figure 2 reveals that both photoperiod (F3,660 = 5.2, P < 0.01) and hibernation duration (F4,660 = 2.6, P < 0.05) showed significant effects on the foundation of colonies, but their interaction effect was insignificant (F12,660 = 0.3, P = 0.99). Significantly higher rates of colonies (67.6%) were produced in LD 8:16 conditions with 3.0 month hibernated queens. The lowest rates (29.4%) were observed in the LD 24:0 treatment of the queens hibernated for 2.0 months.

Rate (%) of Bombus terrestris queens founding colonies according to photoperiodic regimes and hibernation durations (n = 34 queens). Bars with no common letter are significantly different (P ≤ 0.05, Duncan's multiple range test). Photoperiods ( ) 0 h light : 24 h dark (LD 0:24), (
) 0 h light : 24 h dark (LD 0:24), ( ) LD 8:16, (
) LD 8:16, ( ) 16:8 and (
) 16:8 and ( ) 24:0.
) 24:0.
Colony initiation and foundation period
Table 1 shows a great variation of colony initiation periods in response to photoperiods (F3,12 = 15.3, P < 0.001) and hibernation durations (F4,12 = 9.7, P < 0.001). Colonies initiated significantly early (3.4 ± 0.4 and 3.6 ± 0.6 days, respectively) in the LD 8:16 treatment where the queens experienced hibernation durations of 3.5 and 4.0 months, whereas the most significantly delayed periods (11.1 ± 1.8 day) were found in the LD 0:24 treatment where the queens experienced hibernation duration for 2.0 months.
| Photoperiod light : dark (h) | Hibernation duration (month) | ||||
|---|---|---|---|---|---|
| 2.0 | 2.5 | 3.0 | 3.5 | 4.0 | |
| 0:24 | 11.1 ± 1.8 a | 9.1 ± 1.3 ac | 7.4 ± 1.1 bf | 5.6 ± 0.8 dg | 5.1 ± 0.8 dg |
| (24) | (25) | (27) | (28) | (28) | |
| 8:16 | 7.0 ± 0.9 bf | 5.9 ± 0.6 cg | 4.7 ± 0.6 fg | 3.4 ± 0.4 g | 3.6 ± 0.6 g |
| (25) | (29) | (30) | (26) | (26) | |
| 16:8 | 8.1 ± 1.2 ac | 6.4 ± 0.7 bg | 4.9 ± 0.6 eg | 5.0 ± 0.8 dg | 5.6 ± 1.0 dg |
| (23) | (27) | (28) | (26) | (25) | |
| 24:0 | 9.4 ± 1.2 ab | 9.0 ± 1.2 ac | 7.4 ± 0.9 bf | 8.4 ± 1.4 ac | 8.3 ± 1.3 ac |
| (22) | (23) | (27) | (23) | (21) | |
- Means followed by different letters are significantly different (P ≤ 0.05, Duncan's multiple range test). Data expressed as mean days ± SE (n).
Table 2 indicates that the colony foundation period (28.2 ± 0.8 days) was shortest in the LD 8:16 photoperiod belonging to the queens hibernated 3.0 for months. The longest period (32.6 ± 1.3 day) was entrained under an LD 24:0 photoperiod with 2.0 month hibernated queens. An anova of the results showed significant differences (photoperiod: F3,12 = 9.6, P < 0.01; hibernation duration: F4,12 = 6.0, P < 0.01).
| Photoperiod light : dark (h) | Hibernation duration (month) | ||||
|---|---|---|---|---|---|
| 2.0 | 2.5 | 3.0 | 3.5 | 4.0 | |
| L0:D24 | 32.2 ± 1.1 ac | 31.3 ± 1.1 ac | 29.9 ± 1.1 ac | 29.4 ± 1.0 ac | 29.4 ± 1.2 ac |
| (12) | (13) | (15) | (15) | (17) | |
| L8:D16 | 30.4 ± 1.1 ac | 28.9 ± 0.8 bc | 28.2 ± 0.8 c | 28.7 ± 0.8 bc | 28.9 ± 0.9 bc |
| (14) | (16) | (23) | (22) | (19) | |
| L16:D8 | 28.8 ± 1.1 ac | 29.6 ± 1.3 ac | 28.6 ± 1.0 bc | 29.2 ± 0.9 ac | 29.3 ± 0.7 ac |
| (14) | (14) | (20) | (18) | (15) | |
| L24:D0 | 32.6 ± 1.3 a | 30.2 ± 1.2 ab | 29.3 ± 1.2 ac | 31.3 ± 0.8 ac | 31.5 ± 1.4 ac |
| (10) | (11) | (15) | (12) | (11) | |
- Means followed by different letters are significantly different (P ≤ 0.05, Duncan's multiple range test). Data expressed as mean days ± SE (n).
Number of workers from the first brood egg cells
Figure 3 shows a significant effect of photoperiods (F3,12 = 7.9, P < 0.01) on the production of the number of workers of the first brood cells, but not the hibernation durations (F4,12 = 3.0, P > 0.05). In LD 8:16 conditions, 3.5 month hibernated queens produced a significantly higher number of workers (10.2 ± 0.7). On the other hand, queens hibernated for 4.0 months and reared in a LD 24:0 conditions produced the lowest number of workers (7.6 ± 0.9).

Number of Bombus terrestris workers produced from the first brood egg cells in relation to photoperiodic regimes and hibernation durations. Data are expressed as mean ± SE (n = number of queens producing workers). 0 h Light : 24 h dark (LD 0:24): 12, 13, 15, 15 and 17 for 2.0, 2.5, 3.0. 3.5 and 4.0 months, respectively; LD 8:16: 16, 16, 23, 22 and 19 for 2.0, 2.5, 3.0. 3.5 and 4.0 months, respectively; L:D 16:8: 14, 14, 20, 18 and 15 for 2.0, 2.5, 3.0. 3.5 and 4.0 months, respectively; LD 24:0: 10, 11, 15, 12 and 11 for 2.0, 2.5, 3.0. 3.5 and 4.0 months, respectively). Bars with no common letter are significantly different (P ≤ 0.05, Duncan's multiple range test). Photoperiods ( ) LD 0:24, (
) LD 0:24, ( ) LD 8:16, (
) LD 8:16, ( ) LD 16:8 and (
) LD 16:8 and ( ) LD 24:0.
) LD 24:0.
Proportion of queens producing sexual queens
Queens produced progeny queens variably in relation to photoperiod and hibernation duration (Fig. 4). The highest percentage (38.2%) of queens producing new queens was found in LD 8:16 conditions when the colonies were reared with the queens experiencing hibernation durations of 3.0 and 3.5 months. However, the queens reared in LD 24:0 and hibernated for 4.0 months gave the lowest rate (8.8%) of queen production. An anova of the data indicated that photoperiodic regimes have a significant effect on queen production rate (F3,660 = 6.5, P < 0.01), but that hibernation duration (F4,660 = 1.6, P = 0.18) and interaction effects (F12,660 = 0.5, P = 0.93) were insignificant.

Rate (%) of queens of Bombus terrestris produced young queens in relation to photoperiodic regimes and hibernation duration (n = 34 queens). Bars with no common letter are significantly different (P ≤ 0.05, Duncan's multiple range test). Photoperiods ( ) 0 h light : 24 h dark (LD 0:24), (▪) 8:16, (
) 0 h light : 24 h dark (LD 0:24), (▪) 8:16, ( ) 16:8 and (
) 16:8 and ( ) 24:0.
) 24:0.
Production of sexuals
Photoperiods (F3,12 = 62.7, P < 0.001) and hibernation durations (F4,12 = 5.8, P < 0.01) showed significant effects on the production of sexual males. The maximum number of males (171.2 ± 12.2) was produced in the colonies when the queens hibernated for 3.5 months and were reared in an LD 8:16 photoperiod (Table 3). Statistically identical numbers of males (162.5 ± 15.5 and 161.1 ± 12.1) were produced in the photoperiods LD 8:16 and 16:8 respectively, when the queens hibernated for 3.0 months. The minimum number of males (67.2 ± 6.5) was produced in the colonies reared in LD 24:0 conditions when the queens hibernated for 4.0 months. Table 4 shows that progeny queen production was dependent on photoperiod (F3,12 = 35.1, P < 0.0001) but not on hibernation duration (F4,12 = 3.1, P > 0.05). The queens which hibernated for 3.0 months and developed colonies in LD 8:16 conditions produced the highest number of queens (91.2 ± 9.9). On the other hand, the lowest number of queens (20.3 ± 8.9) were produced in an LD 24:0 photoperiod when the queens experienced hibernation duration for 4.0 months.
| Photoperiod light : dark (h) | Hibernation duration (month) | ||||
|---|---|---|---|---|---|
| 2.0 | 2.5 | 3.0 | 3.5 | 4.0 | |
| 0:24 | 111.5 ± 12.5 ad | 117.8 ± 11.2 ad | 143.4 ± 18.8 ac | 142.8 ± 21.9 ad | 133.4 ± 15.9 ad |
| (9) | (10) | (10) | (13) | (14) | |
| 8:16 | 118.9 ± 13.2 ad | 139.3 ± 16.3 ad | 162.5 ± 15.5 a | 171.2 ± 12.2 a | 147.2 ± 11.7 ab |
| (10) | (11) | (19) | (14) | (14) | |
| 16:8 | 129.1 ± 10.5 ad | 136.2 ± 16.6 ad | 161.1 ± 12.1 a | 145.4 ± 12.0 ac | 137.2 ± 11.1 ad |
| (8) | (10) | (13) | (13) | (12) | |
| 24:0 | 72.4 ± 12.5 bd | 78.5 ± 9.3 bd | 80.0 ± 8.5 bd | 70.7 ± 8.2 cd | 67.2 ± 6.5 d |
| (6) | (8) | (8) | (7) | (7) | |
- Means followed by different letters are significantly different (P ≤ 0.05, Tukey HSD post hoc test). Data expressed as mean number ± SE (n).
| Photoperiod light : dark (h) | Hibernation duration (month) | ||||
|---|---|---|---|---|---|
| 2.0 | 2.5 | 3.0 | 3.5 | 4.0 | |
| 0:24 | 49.2 ± 10.5 ab | 54.3 ± 9.3 ab | 58.4 ± 13.1 ab | 66.3 ± 16.0 ab | 69.2 ± 18.4 ab |
| (5) | (6) | (7) | (6) | (5) | |
| 8:16 | 58.2 ± 8.8 ab | 67.9 ± 9.4 ab | 91.2 ± 9.9 a | 80.3 ± 9.5 ab | 65.9 ± 6.2 ab |
| (6) | (7) | (13) | (13) | (12) | |
| 16:8 | 53.4 ± 9.2 ab | 56.6 ± 9.6 ab | 77.4 ± 11.7 ab | 71.9 ± 14.8 ab | 60.9 ± 9.1 ab |
| (5) | (7) | (10) | (8) | (8) | |
| 24:0 | 25.3 ± 6.7 b | 32.5 ± 7.3 ab | 28.0 ± 6.8 b | 24.8 ± 9.1 b | 20.3 ± 8.9 b |
| (4) | (4) | (5) | (4) | (3) | |
- Means followed by different letters are significantly different (P ≤ 0.05, Tukey HSD post hoc test). Data expressed as mean number ± SE (n).
DISCUSSION
Photoperiod is one of the main cues regulating the diapause of insects (Hodek & Ruzicka 1979). In the present study, artificial photoperiods showed significant effects on the colony development of hibernated B. terrestris queens. The results reveal that the highest rate of queens (88.2%) initiating colonies were found in LD 8:16 conditions when the queens experienced hibernation for 3.0 months.
Colony initiation period was shortest (3.4 ± 0.4 and 3.6 ± 0.6 days) in the photoperiodic treatment of LD 8:16 when queens hibernated for 3.0 and 3.5 months. Our results are in agreement with Tasei (1994) who reared narcotized B. terrestris queens under fluorescent tubes at LD 8:16 and constant dark conditions, and observed that mean delay in egg laying was significantly reduced at LD 8:16 compared to LD 0:24. He also found that in light : dark conditions, 97% queens laid their first eggs within 40 days, whereas in the dark condition oviposition rates were only 59%. Yoon and Kim (2003) suggested that oviposition rates of B. ignitus (Bombus ignitus) Smith queens were higher in LD 8:16 and 16:8 than in 0:24 conditions. Overwintered B. terrestris queens showed higher rates of oviposition under artificial short day (LD 8:16) conditions as compared to LD 0:24 and 16:8 conditions (Yoon et al. 2003). They also mentioned that preoviposition period was shorter in the same photoperiodic regime.
Artificial photoperiods and hibernation durations showed remarkable effects on colony foundation. In this study, colony foundation rate was significantly highest (67.6%) in the queens experiencing hibernation duration for 3.0 months and reared at a photoperiod of LD 8:16. This might be due to a higher rate of oviposition, early initiation of the colony and direct influence of the queen on the first brood cells in a suitable photoperiodic environment.
Significantly the shortest period (28.2 ± 0.8 days) of colony foundation was observed in the queens hibernated for 3.0 months and kept at an LD 8:16 photoperiod. This artificial short day is in concordance to the natural day length of the temperate region in spring, when bumblebees initiate colonies (Tasei 1994). Hatching of eggs is an indicating factor of colony foundation that is affected by photoperiods. In the corn stalk borer, Sesamia nonagrioides Lef, egg hatching was higher under LD 10:14 compared to 16:8 (Fantinou et al. 2004).
For successful colony development, early colony initiation and the emergence of enough workers in the first brood is a prerequisite. Pomeroy and Plowright (1982) studied the relationship between worker numbers and the production of sexuals in B. perplexus (Bombus terrestris) Cresson, and reported that a significantly greater number of queens appeared in the colonies that had higher numbers of workers in the first brood. Reproductive success of B. ruderatus (Bombus ruderatus) Fab. was strongly influenced by the peak size of the worker population in the colony (Pomeroy 1979). The present findings show that a higher number of workers in the first brood was found when the queens hibernated for 3.5 months and were reared under LD 8:16 conditions.
The rates of founding queens producing new queens were significantly highest (38.2%) in the photoperiod LD 8:16 with a hibernation duration of 3.0 and 3.5 months. In the same photoperiodic treatment, 3.0 month hibernated queens produced significantly higher numbers (91.2 ± 9.9) of young queens. These data are in agreement with Yoon and Kim (2003), who reported that the queen production rate of B. ignitus increased twofold in LD 8:16 or 16:8 compared to 0:24 conditions. Hodek and Iperti (1983) observed twice the fecundity of the coccinellid Semiadalia undecimnotata Schneider under short day compared to long day conditions.
Sexual male production was also significantly higher (171.2 ± 12.2 and 162.5 ± 15.5) in an LD 8:16 photoperiod when the queens had experienced hibernation for 3.5 and 3.0 months. Statistically identical numbers (161.1 ± 12.1) of males were produced in an LD 16:8 photoperiod when the queens hibernated for 3.0 months. So, the results indicate that exposure of hibernated bumblebee queens to artificial light influenced the production of sexuals. Lees (1964) studied the sexual and parthenogenetic daughters of Megora under typical long and short day conditions and stated that production was controlled by the direct action of illumination on the brain of the parent.
The present findings show that the rate and period of colony initiation, colony foundation, number of workers of the first brood egg cells and total number of sexuals of B. terrestris vary with response to photoperiods and hibernation durations. This was suggested by Grinfeld and Zacharova (1971), who stated that bumblebees have a clear photoperiodic reaction. They demonstrated that artificial shortening of day length reduced the development cycle of Bombus hortorum L and Bombus agrorum Calam, and triggered the production of sexuals. However, the results so far indicate that the queens reared in an LD 8:16 photoperiod which had experienced hibernation duration for 3.0 months had a greater capacity for colony development. Additionally, the intensities and wavelengths of light are important parameters for reproduction and development. Therefore, further research is necessary to improve the lighting system with regards to spectrum and intensity.

