

Impacts of Hurricane Frances on Florida scrub-oak ecosystem processes: defoliation, net CO2 exchange and interactions with elevated CO2
Abstract
Hurricane disturbances have profound impacts on ecosystem structure and function, yet their effects on ecosystem CO2 exchange have not been reported. In September 2004, our research site on a fire-regenerated scrub-oak ecosystem in central Florida was struck by Hurricane Frances with sustained winds of 113 km h−1 and wind gusts as high as 152 km h−1. We quantified the hurricane damage on this ecosystem resulting from defoliation: we measured net ecosystem CO2 exchange, the damage and recovery of leaf area, and determined whether growth in elevated carbon dioxide concentration in the atmosphere (Ca) altered this disturbance. The hurricane decreased leaf area index (LAI) by 21%, which was equal to 60% of seasonal variation in canopy growth during the previous 3 years, but stem damage was negligible. The reduction in LAI led to a 22% decline in gross primary production (GPP) and a 25% decline in ecosystem respiration (Re). The compensatory declines in GPP and Re resulted in no significant change in net ecosystem production (NEP). Refoliation began within a month after the hurricane, although this period was out of phase with the regular foliation period, and recovered 20% of the defoliation loss within 2.5 months. Full recovery of LAI, ecosystem CO2 assimilation, and ecosystem respiration did not occur until the next growing season. Plants exposed to elevated Ca did not sustain greater damage, nor did they recover faster than plants grown under ambient Ca. Thus, our results indicate that hurricanes capable of causing significant defoliation with negligible damage to stems have negligible effects on NEP under current or future CO2-enriched environment.
Introduction
On average, a hurricane with an intensity of Saffir–Simpson Scale 3–5 makes landfall along the eastern US coastline 2 out of 3 years (Smith, 1999; McNulty, 2002). Hurricane disturbances have profound long-term effects on the species composition, structure and succession in many ecosystems (Tanner et al., 1991; Foster et al., 1998; Turner et al., 2003). In the short-term, hurricanes cause defoliation (McNulty, 2002) and alter nutrient cycling (Erickson & Ayala, 2004). The impacts of hurricanes on ecosystem CO2 exchange have not been reported.
Defoliation is the most common damage from catastrophic wind disturbance (Everham & Brokaw, 1996; Ostertag et al., 2003). Most plants produce new leaves, even out of regular foliation period, to compensate foliage loss (Boucher et al., 1990; Basnet, 1993; Everham & Brokaw, 1996). Factors affecting leaf area index (LAI), the amount of leaf area per unit ground area, are likely to alter canopy photosynthesis through light interception (Cowling & Field, 2003; Ewert, 2004) and modify respiration through control on substrate supply (Högberg et al., 2001; Whitehead et al., 2004; Tang et al., 2005; Hartley et al., 2006; Trumbore, 2006). As carbon dioxide concentration in the atmosphere (Ca) increases (Prentice et al., 2001), it is not known whether the impacts of hurricanes on defoliation, refoliation, and ecosystem CO2 exchange will be altered. A positive answer to this question could be postulated based on evidence that elevated Ca can increase photosynthesis and LAI (Hymus et al., 2002; Dermody et al., 2006) and that the additional photosynthate would help plants in refoliation, as has been documented after simulated defoliation from herbivory (Fajer et al., 1991; Kruger et al., 1998). On the other hand, a negative answer can also be suggested based on reports that elevated Ca often has little effect on LAI (Drake et al., 1997; Cowling & Field, 2003) and other reports showing that refoliation was not stimulated by elevated CO2 after simulated defoliation from herbivory (Lovelock et al., 1999; Volin et al., 2002). To our knowledge, no study has explored the combined impacts of hurricane disturbance and elevated Ca on ecosystem processes, at least in part, because such large-scale natural disturbances are remarkably difficult to anticipate and include in experimental designs (Everham & Brokaw, 1996).
We have been using eddy covariance since 2000 to continuously measure carbon dioxide exchange between a fire-regenerated scrub-oak ecosystem and the atmosphere at our Ameriflux research site on the Merritt Island National Wildlife Refuge, Cape Canaveral, FL, USA (Dore et al., 2003). Concurrently, we have been studying the effects of elevated Ca using 16 open-top chambers (eight at normal ambient and eight at normal ambient + 350 μmol mol−1) since 1996. In September 2004, our research site was struck by Hurricane Frances with sustained winds of 113 km h−1 and wind gusts as high as 152 km h−1. This provided a unique opportunity for studying the impacts of a hurricane on this ecosystem under current conditions and elevated Ca. The objectives of this study were: (i) to investigate the hurricane damage on leaf area and ecosystem CO2 exchange; (ii) to describe the recovery from the damage; and (iii) to determine whether growth in elevated Ca altered this disturbance and rate of recovery.
Materials and methods
Experimental site
The experimental site was located on the Merritt Island National Wildlife Refuge, Cape Canaveral, FL, USA (28°38′N, 80°42′W), in a warm, humid subtropical climate. Annual precipitation averages 131 cm, with a dry period typically occurring from April to June. The mean daily maximum temperature is 22.3 °C for January and 33.3 °C for July, and the mean daily minimum temperature is 9.6 °C for January and 21.9 °C for July (Mailander, 1990). Thunderstorms are common in the summer with frequent lightning strikes, which often cause wildfires.
The soil consists primarily of sand and sandy coquina, deposited since the Pleistocene, and has a 2–5 cm O horizon and 10–20 cm A horizon. The composition of aboveground biomass at the research site was Quercus myrtifolia Willd. (76%), Q. geminata Small (15%), and Q. chapmanii Sarg (7%). Additional species included Myrica cerifera L., Lyonia ferruginea (Walt.) Nutt, and Galactia elliottii Nuttall (Dijkstra et al., 2002). This ecosystem is nutrient limited, naturally fire dependent, and managed by controlled burns (Schmalzer & Hinkle, 1992). The scrub-oak canopy was 34–52 cm in 1998 and 90–150 cm in 2005.
Before the study, aboveground biomass was measured and the site was burned in January 1996. After burning, plots were assigned to blocks of three plots with similar pre-burn biomass characteristics. Sixteen open-top chambers (eight at ambient and eight at 350 μmol mol−1 above ambient Ca) were erected over burned plots, with eight additional unchambered plots that served as chamber controls. The chambers were octagonal, 3.6 m in diameter, and 2.1 m in height having an area of 9.45 m2. Pure CO2 was added to the air stream blown into the elevated treatment chambers. Shoots of plants that had begun to grow after the site had been burned were removed before beginning treatment with elevated Ca on May 14, 1996. Elevated Ca (350 μmol mol−1 above ambient) treatments were monitored continuously, 24 h a day. Further details about the experimental site were reported by Dijkstra et al. (2002).
In 2000, an eddy covariance station was established in the same stand as the chambers approximately 100 m NW of the closest chamber. The anemometer and CO2 analyzer were mounted on top of a mast placing the sensor head at 3.5 m, high enough to measure in the internal boundary layer and to avoid disturbance associated with canopy roughness. Footprint analyses revealed that the area of major contribution to the measured flux was between 15 and 30 m from the tower. Measurements were not affected by the CO2 emission from the chambers maintained at elevated Ca (Dore et al., 2003).
Hurricanes
Our research site has been visited by several hurricanes since May 1996. On September 15 and October 16, 1999, Hurricanes Floyd and Irene with sustained winds of 113 and 121 km h−1, respectively, impacted our site and caused some defoliation (not quantified). On August 13, 2004, Hurricane Charley crossed our research site with sustained winds of 54 km h−1 and gusts of 76 km h−1 and caused negligible defoliation because of its low wind speed and fast movement (33 km h−1). On September 5, 2004, the eye of Hurricane Frances made landfall near Sewall's Point on the east coast of Florida, about 160 km south of our research site. The sustained wind speed at landfall was 169 km h−1, which made this a category II hurricane. The storm crossed Florida from southeast to northwest at a speed of 13 km h−1 over 3 days. Our experimental site experienced sustained winds of 113 km h−1 and gusts as high as 152 km h−1, which is characteristic of a category I hurricane. Twenty days later, Hurricane Jeanne hit the experimental site with a much lower wind speed of 58 km h−1 and caused negligible damage. The hurricane disturbance discussed in this report was from Hurricane Frances.
On September 2, 2004, 3 days before Hurricane Frances, all the chamber walls were removed and CO2 enrichment stopped. Chambered plants were exposed to the same winds as those outside the chambers. After Hurricane Jeanne, chamber walls were reinstalled and CO2 enrichment resumed on October 1, 2004. We describe the disturbance of Hurricane Frances in terms of leaf area, litter fall, and ecosystem CO2 exchange measured using eddy flux and using the open top chambers as cuvettes.
Leaf fall
Before the hurricane, leaf fall was collected monthly from sixteen 0.1 m2 traps that were situated every 10 m along four randomly oriented transects radiating away from the eddy covariance system. Samples were dried for three days at 70 °C and then weighed.
LAI
LAI was measured monthly before and after the hurricane by applying Beer's Law to the attenuation of photosynthetic photon flux density (PPFD) through the canopy using a 40 cm long ceptometer containing 40 individual sensors (SF-40, Decagon, Pullman, WA, USA). The details on the validation of this method for use within the open top chambers have been reported by Hymus et al. (2002). At the eddy flux site, the measurements were carried out at 20 points 5 m apart along a N–S or E–W transects with the tower at the mid-point of the two transects.
Refoliation measurements
We measured refoliation on the seven largest individuals in each plot of the two dominant oak species Q. myrtifolia and Q. geminata, plants which had been marked and monitored since April 2004. Large plants were chosen because the largest 5% of plants per chamber represented 60% of the total biomass and produced 60% of biomass increment during 2004. After the hurricane, we began monitoring the total number of leaves of the marked plants, counting leaves on October 6–9, October 26–29, November 16–19, 2004 and March 10–15, 2005. During each of these periods, we also measured basal stem diameters of the marked plants to determine their biomass (Dijkstra et al., 2002), necessary for scaling leaf area recovery from marked plants to a ground area basis.
We estimated leaf area on the marked plants as the product of the number of new leaves and average leaf area per leaf (Nobel & Long, 1985; Eschenbach & Kappen, 1996). To determine average leaf area per leaf, two new branches from the top, middle, and bottom of the canopies of the marked Q. myrtifolia and Q. geminata plants were sampled from each chamber following leaf counting. Total number of leaves in each of the two branches was counted and leaf area measured separately with an area meter (LI-COR, Model 3100, Lincoln, NE, USA). These calculations were carried out separately for each treatment, although there was no statistically significant difference in the average leaf area per leaf between the treatments (data not shown; Hymus et al., 2002).
We scaled leaf area recovery on these marked plants to recovery of total plot leaf area using leaf area: biomass relationships and measurements of total plant biomass in each plot. First, we calculated new leaf area per unit of aboveground biomass for each marked plant. New leaf area per unit ground area (total plot leaf area recovery) was estimated as the product of leaf area recovery per unit biomass of the marked plants and total plot biomass per unit ground area. Aboveground plant biomass has been measured each year since the beginning of the CO2 enrichment in 1996 (Dijkstra et al., 2002).
Measurement of net ecosystem CO2 exchange (NEE) by eddy covariance
Eddy covariance was used to measure NEE from January 2002 to March 2006 (Aubinet et al., 2000; Baldocchi, 2003; Dore et al., 2003). A closed-path system was used between January 2002 and March 2004 and further details were reported by Dore et al. (2003). An open-path system was used between March 2004 and March 2006 and consisted of a CSAT3 sonic anemometer (Campbell Scientific, Logan, UT, USA) and a LI-7500 infrared gas analyzer (LI-COR). The LI-7500 head was situated 20 cm horizontally from the sonic anemometer head and tilted 45°N to prevent contamination from solar radiation. A comparison of data collected simultaneously from both systems showed that there was only a 3% difference in CO2 flux measurements (NEEclose= 1.03 NEEopen+ 0.49, R= 0.98**, n= 1021). Raw data were logged at 10 Hz and the WPL correction (Webb et al., 1980), coordinate rotation, and frequency response correction were applied to mean half-hourly fluxes of NEE. The degree of energy closure was > 85% for each system each year. In a comparison with simultaneous measurements from the roving Ameriflux calibration system, deviation was less than 3%.
 (1)
(1) (2)
(2)Measurement of NEE by chamber
The chamber method of measuring NEE was described in detail by Dore et al. (2003) and Hymus et al. (2003). In brief, open-top chambers were used as gas-exchange cuvettes. When the measurements were conducted, perforated lids were added to prevent backflow of ambient air into the chamber. NEE was determined by multiplying the flow rate of air through the open-top chambers by the difference in CO2 concentration between air coming into and air leaving the chambers. NEE measured using this method was not different from simultaneous measurements using an eddy covariance system (Dore et al., 2003).
NEE was measured in the chambers once a month before the hurricane since 1997. The measurements of NEE were made only in two ambient and two elevated CO2 chambers because the damage to the equipment required for NEE measurements could not be immediately repaired for all the chambers. Light-response curves were fit to a rectangular hyperbola as were done for the above-mentioned eddy covariance study. Using monthly light-response curves, NEE at PPFD of 1500 μmol m−2 s−1 was defined as NEEmax for measurements inside the chambers.
Soil respiration
Soil respiration (Rs) was measured monthly with a LI-6400-09 portable gas exchange system (LI-COR). Measurements were taken from one fixed plot (soil area exposed: 72 cm2 per plot) per chamber since 1999 and from 22 fixed plots (soil area exposed: 72 cm2 per plot) within the eddy covariance footprint since June 2000. Simultaneous measurements of soil respiration and soil temperature (measured adjacent to the soil respiration plots) were fit to Eqn (2) to evaluate the relationship between both variables.
Meteorological measurements
PPFD (LI-190, LI-COR), air temperature (copper/constantan thermocouple, Omega, Stamford, CT, USA), relative humidity (HMP45C, Campbell Scientific), and precipitation (TE525 tipping bucket, Campbell Scientific) were measured continuously on a 3.5 m meteorological tower located 30 m from the eddy flux tower. Soil temperature (copper/constantan thermocouple, at 10 cm depth) and soil water content (CS615, Campbell Scientific, integrated over the top 15 cm of soil) were measured adjacent to the meteorological tower and in the ambient and elevated Ca chambers.
Statistics
Results of leaf fall and LAI from eddy covariance measurements (Fig. 1) were analyzed with systat 11 software package (Systat Software Inc., Richmond, CA, USA, 2004) using repeated measures analysis of variance (anova), where year was considered the between-subjects contrast and month was the repeated measure (within-subjects contrast). Foliage damage from chamber measurements (Fig. 5) were analyzed using two-way repeated measures anova, where year and CO2 treatment were considered two independent between-subjects contrasts and month was the repeated measure (within-subjects contrast). Results of foliage recovery from chamber measurements (Fig. 6) were analyzed using repeated measures anova, where CO2 treatments were considered between-subjects contrast and time after hurricane was the repeated measure (within-subjects contrast). LAI, NEEmax, soil respiration, soil temperature, and soil water content from chamber measurements (Fig. 7) were analyzed using repeated measures anova, where CO2 treatment was considered between-subjects contrast and year was the repeated measure (within-subjects contrast). For all the repeated anova, when a significant effect was found, a post hoc test for repeated measures was further conducted to detect exactly which means were significantly different, where the experiment-wise error rate was set to 0.05 using Bonferroni Correction for multiple comparisons. GPP, Re, and NEP from eddy covariance measurements (Fig. 3) have no replicate, so repeated anova is not feasible. They were analyzed using paired t-test, where data from the same month were considered a pair. Parameters from regressions between NEEday and PPFD and between soil respiration and soil temperature (Fig. 2) were analyzed using dummy variables with error terms generated by sigmaplot 8.0 to determine whether the coefficients for different regression lines were significantly different (Gujarati, 1970; Powell et al., 2005).

Leaf fall (means±SE, n= 16) and leaf area index (means±SE, n= 20) at the eddy flux site in 2003–2005.

Leaf area index at ambient (open symbols) and elevated (filled symbols) CO2 in 2003–2005. Data are means±SE (n= 8).

Post-hurricane refoliation at ambient (open circles) and elevated (filled circles) CO2. Data are means±SE (n= 8).

Leaf area index, maximum daytime net ecosystem CO2 exchange (NEEmax), soil respiration, soil temperature at 10 cm (circles) and soil water content between 0 and 15 cm (squares) in Octobers of 2001–2005 at ambient (open symbols) and elevated (closed symbols) CO2. Data are means + or ± SE (n= 2 for NEEmax; n= 8 for leaf area index, soil respiration, soil water content, and soil temperature). Soil temperature is not available due to equipment malfunction in October 2002.

Monthly gross primary production (GPP), ecosystem respiration (Re), and net ecosystem production (NEP) at the eddy flux site in 2002–2005.

Daytime net CO2 exchange (NEEday) and soil respiration as a function of photosynthetic photon flux density (PPFD) and soil temperature at 10 cm, respectively, before (open squares, dash lines), immediately after (filled squares, solid lines), and 1 year after (open triangles, dotted lines) the hurricane disturbance. For brevity, only 10% of the data were randomly chosen and represented to indicate the variability of the data. Data were from eddy covariance measurements as described in Table 1. All the regressions are statistically significant (P < 0.01).
Results
Eddy covariance: leaf fall and LAI
In a typical year, such as 2003, scrub-oak species have a dominant pulse of leaf fall in March when new foliage starts to emerge (Fig. 1a). In 2004, the hurricane caused a premature leaf fall pulse in September as indicated by repeated measures anova comparing 2004 with 2003 (P= 0.028) and post hoc Bonferroni test for repeated measures (P < 0.05). In 2005, leaf fall followed the same path as in 2003; repeated measures anova revealed no difference from 2003 (P= 0.838).
No significant difference was found in LAI from January to August between 2003 and 2004 (repeated measures anova, P= 0.691). The hurricane abruptly reduced LAI in September by 21% from the August maximum value of 2.3 (P= 0.002) (Fig. 1b). In 2005, LAI followed the same path as in 2003; repeated anova revealed no difference from 2003 (P= 0.722).
Eddy covariance: ecosystem CO2 exchange
In October 2004, one month after the hurricane, daytime NEE at saturating light [parameter NEEsat in Eqn (1)] was significantly reduced by 16% (P < 0.001) compared with that in October 2002 and 2003 (Fig. 2a). Before the hurricane, in July and August 2004, the response of daytime NEE (NEEday) to PPFD was similar to that of the same months in the previous 2 years (data not shown). In October 2005, 1 year after the hurricane, NEEday was not different from that in Octobers 2002 and 2003 (P= 0.346).
The sensitivity of soil respiration to soil temperature was evaluated by comparing model parameter b [Eqn (2)] from the pre-hurricane, hurricane, and post-hurricane periods. The sensitivity of soil respiration to soil temperature declined by 46% (P= 0.006) after the hurricane (Fig. 2b). The sensitivity recovered to pre-hurricane level in 2005 (P= 0.684).
Minimum values of GPP occurred between December and February, and maximum values between May and September (Fig. 3). The hurricane reduced GPP and Re by 22% (P= 0.067) and 25% (P= 0.020), respectively, on average from September through November compared with the average values in 2002 and 2003. There was no significant difference in GPP and Re during all other months of 2004 compared with the average values in 2002 and 2003. In 2005, GPP and Re were 11% (P= 0.016) and 25% (P= 0.04) lower in January through June compared with average values in 2002 and 2003, indicating that they were not recovered yet from the hurricane damage. In July through October 2005, GPP and Re were the same as in the same period of 2002 and 2003, indicating they were fully recovered. NEP in 2004 was not different from the average values in 2002 and 2003. NEP in 2005 was also not different from the average values in 2002 and 2003. Re was positively correlated with GPP in the measuring periods (Fig. 4).

Correlation of monthly ecosystem respiration (Re) with monthly gross primary production (GPP) at the eddy flux site in 2002–2005 (2002: Re= 0.87 GPP + 1.37, R= 0.97**; 2003: Re= 0.75 GPP + 11.63, R= 0.91**; 2004: Re= 0.92 GPP − 10.07, R= 0.96**; 2005: Re= 0.99 GPP − 36.73, R= 0.91**).
Chamber: defoliation and post-hurricane refoliation
No difference in LAI was found in August between 2003 and 2004 at ambient or elevated Ca (Fig. 5). The hurricane reduced LAI by 41% (P= 0.001) and 48% (P= 0.001) at ambient and elevated Ca, respectively. The difference in the relative reduction of LAI between the CO2 treatments was not statistically significant. Elevated Ca increased LAI by 46% (P= 0.014) in 2003 and 2004. After the hurricane, LAI was still higher (30%, P= 0.033) at elevated than at ambient Ca. In 2005, LAI did not differ from that in 2003 at ambient or elevated Ca in August through December (P > 0.217).
New leaves started to emerge within a month following defoliation from the hurricane, well before the typical leaf-out period of April to July. New leaf area generated from refoliation reached a plateau after about 2.5 months (Fig. 6a). Elevated Ca increased the new leaf area per ground area by 104% with marginal significance (P= 0.09). Relative leaf area from refoliation compared with pre-hurricane values was not affected by elevated Ca (Fig. 6b, P= 0.273), being about 10% of pre-hurricane values and compensating for about 20% of the defoliation loss at both ambient and elevated Ca.
Chamber: ecosystem CO2 exchange
The hurricane reduced LAI by 36% and soil respiration by 26% compared with previous years at ambient or elevated Ca (2004 vs. 2001–2003, P < 0.003 for LAI, P < 0.02 for soil respiration, post hoc Bonferroni test, Fig. 7a). Elevated Ca increased LAI by 37% (P= 0.004) and soil respiration by 32% (P= 0.049) on average. In October 2005, 1 year after the hurricane, LAI and soil respiration were about the same as the averages of the 3 years before the hurricane (2004 vs. 2001–2003, P= 0.279 for LAI, P= 0.33 for soil respiration, post hoc Bonferroni test). NEEmax in 2004 was 42% lower than the previous 3 years on average with a significant difference from 2003 (2004 vs. 2003, P= 0.034, post hoc Bonferroni test, Fig. 7b). In October 2005, soil respiration was not different from the average of the three years before the hurricane (2005 vs. 2001–2003, P= 0.113, post hoc Bonferroni test, Fig. 7c). Overall, elevated Ca increased NEEmax by about 30% (P= 0.057). When pooling all the data from Octobers of 2001–2004, NEEmax and soil respiration (Rs) were linearly correlated with each other and with LAI, and CO2 treatments appeared to fall on the same relationships (Fig. 8).

Correlations between soil respiration (Rs) and maximum daytime net ecosystem CO2 exchange (NEEmax), NEEmax and leaf area index (LAI), and Rs and LAI, respectively (Rs= 0.11 NEEmax+ 3.25, R= 0.88**; NEEmax= 16.62 LAI + 1.66, R= 0.90**; Rs= 2.56 LAI + 2.47, R= 0.95**). Data were from ambient (open circles) and elevated (filled circles) CO2 in Octobers of 2001–2005 as described in Fig. 7.
Soil water content between 0 and 15 cm was not affected by elevated Ca (P= 0.319) and did not significantly change in the period of the measurements (P= 0.124) (Fig. 7d). Soil temperature at 10 cm varied in a range of < 4 °C. To analyze whether the temperature difference altered the hurricane effect, we first scaled soil respiration values to the soil temperature in 2005 (25 °C at ambient and elevated CO2) using the three regressions in Fig. 2 and Table 1. We did not use the values in 2002 because soil temperature was not measured due to equipment malfunction. After the temperature correction, soil respiration after the hurricane in 2004 was still significantly different from pre-hurricane in 2003 (P < 0.001, post hoc repeated measures Bonferroni test). Compared with 2003, the hurricane reduced soil respiration by 30% and 27% before and after temperature correction, respectively. Without the 2002 temperature data, soil respiration was still linearly correlated with LAI before and after the temperature correction (before: Rs= 2.91 LAI + 1.58, R= 0.97**; after: Rs= 2.78 LAI + 2.44, R= 0.92**). The temperature correction also did not effect the correlation between soil respiration and NEEmax (after the correction without 2002: Rs= 0.12 NEEmax+ 3.83, R= 0.95**). These analyses indicate that the soil temperature variation of < 4 °C in the measurements had very limited influence on our analyses.
| Data | Pre-hurricane | Hurricane | Post-hurricane |
|---|---|---|---|
| NEEday | October 2002 and 2003, pooled | October 2004 | October 2005 |
| Soil respiration | September 2002–March 2003, September 2003–March 2004, pooled | September 2004–March 2005 | September 2005–March 2006 |
Parameters fromregressions between soilrespiration (Rs) andtemperature (Ts):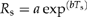 |
|||
| a | 0.6639 | 1.0427 | 0.4555 |
| b | 0.0875 | 0.0574 | 0.1075 |
Discussion
Damage from the hurricane
Fire and hurricanes are the major large-scale disturbances to forest ecosystem in the southeast United States (Myers & Ewel, 1990). Damage to the Florida scrub-oak ecosystem by Hurricane Frances was primarily defoliation as indicated by leaf fall and LAI reduction with no stem breakage or uprooting. This is consistent with reports that defoliation is the most common and wide-spread damage from catastrophic wind disturbance (Brokaw & Walker, 1991; Everham & Brokaw, 1996; Ostertag et al., 2003). The hurricane reduced LAI by 21% compared with pre-hurricane values. The damage was large (reduction in LAI by 0.5), nearly as large as seasonal variation in LAI, which ranged from a maximum of 2.3 to a minimum of 1.6 in 2003, 2004 and 2005.
To our knowledge, this study is the first to document the impacts of a hurricane disturbance on ecosystem CO2 exchange. Our finding that hurricane damage reduced GPP and Re is best explained by the observed reduction in LAI. Hurricane damage to the vegetation did not affect leaf-level photosynthesis measured in the months before and after the hurricane or in new vs. old leaves measured after the hurricane (data not shown). We observed a reduction in soil respiration following the hurricane disturbance, consistent with previous findings following a hurricane in Puerto Rico (Steudler et al., 1991). The reduction in soil respiration accounted for effects on total ecosystem respiration, consistent with past work showing that soil respiration accounts for up to 87% of ecosystem respiration in this (Hymus et al., 2003) and in other forest ecosystems (Lavigne et al., 1997; Davidson et al., 1998; Law et al., 1999; Ekblad & Högberg, 2001). The compensatory reduction in GPP and Re resulted in no significant change in NEP (Fig. 3).
Recovery from the damage
Recovery from hurricane damage depends on the magnitude of damage. Damage from hurricanes can be: (i) defoliation (ii) defoliation plus intensive structural damage to stems. Defoliation is the most prevalent type, occurring even when there is no damage to stems (Brokaw & Walker, 1991; Everham & Brokaw, 1996; Ostertag et al., 2003) and without intensive structural damage, recovery occurs in months to years (Walker, 1991; Ostertag et al., 2003). Recovery from structural damage takes years to decades (Merrens & Peart, 1992; Turner et al., 2003). The damage from Hurricane Frances belongs to the first case, and recovery of LAI (1, 5), GPP, and Re (2, 3) appeared to be complete after a year. Thus, recovery of leaf area from hurricane damage is rapid if the damage is limited to defoliation with negligible stem damage (Walker, 1991; Ostertag et al., 2003).
Interaction with elevated CO2
We found no interactions between hurricane and elevated Ca. Although the hurricane caused more absolute damage to LAI at elevated Ca, the relative reduction in LAI compared with pre-hurricane values did not differ between Ca treatments (Fig. 5). Past work shows that hurricane damage is positively correlated with plant size (Everham & Brokaw, 1996; Harrington et al., 1997; Herbert et al., 1999); our results suggest that this relationship is unaffected by elevated Ca. Elevated Ca has been shown to have no effect on wood density (Ceulemans et al., 2002; Calfapietra et al., 2005) or to slightly increase it (Kostiainen et al., 2004). These results suggest that the vulnerability of trees of comparable size to hurricane damage is unlikely to change as Ca rises.
The rate of recovery from defoliation was also not affected by elevated Ca (Fig. 6). This is consistent with the finding that the relative growth rate of aboveground biomass growth rate was the same at ambient and elevated Ca after canopy closure in 2000 (data not published). Overall mean LAI has not increased since 2000 (Fig. 7a). Fine root growth also reached closure in 1999 (Day et al., 2006). Thus, by 2000, the scrub-oak ecosystem had reached a fully coupled stage as defined by Körner (2006), a stage where the recovery of the scrub-oak ecosystem from hurricane damage is unlikely to benefit from rising Ca.
Nutrient pulses could affect recovery following disturbance, though such pulses are often difficult to detect (Vitousek & Denslow, 1986; Uhl et al., 1998; Ostertag et al., 2003). The scrub-oak ecosystem is nutrient poor (Schmalzer & Hinkle, 1992) and aboveground growth may be sensitive to nutrient pulses. Using the product of live leaf N concentration and defoliation measured here, we estimate that the hurricane transferred 1.63 g N m−2 from live leaves to dead leaves on the forest floor in ambient Ca, and 2.62 g N m−2 in elevated Ca. While this is a substantial N flux compared with annual rates of N uptake from soil at this site (Hungate et al., 2006), the increment caused by elevated Ca, along with the slow rates of N release during decomposition shown at the site (Dilustro et al., 2001), suggests that this mechanism is very unlikely to elicit a differential CO2 response following hurricane disturbance.
Control of photosynthesis on respiration
Globally, ecosystem respiration is one of the largest gross fluxes in the annual carbon budget, 18 times the rate of fossil fuel release in the 1990s (Prentice et al., 2001). Factors affecting the balance between photosynthesis and respiration could potentially influence atmospheric Ca. In this study, the linear relationships between Re and GPP (Fig. 4) and between soil respiration and daytime maximum NEEmax (Fig. 8a) indicate importance of photosynthesis in mediating respiration. These findings are consistent with recent studies revealing that about half of all CO2 released from soil comes from photosynthate produced a few days earlier (Steinmann et al., 2004; Tang et al., 2005). Current assimilate to roots is a key driver of soil respiration (Högberg et al., 2001; Kuzyakov & Cheng, 2001; Hartley et al., 2006). For example, in a Scots pine forest in northern Sweden, soil respiration decreased up to 37% within 5 days and by 56% 2 weeks after girdling and thus terminating the flux of photosynthates from the tree canopy to the roots through the phloem (Högberg et al., 2001). The control of photosynthesis on soil respiration was also confirmed by the EUROFLUX project with a large and intensive study on carbon exchange in European forests (Janssens et al., 2001). On average, 80% of GPP was respired by autotrophs and heterotrophs and released back to the atmosphere. Annual soil respiration was positively and linearly correlated with annual GPP but not with annual temperature (Janssens et al., 2001).
Substrate availability for soil respiration is extremely difficult to measure compared with soil temperature and soil water content because substrates consist of a wide range of compounds and are highly dynamic (Davidson et al., 2006). LAI has been suggested as a surrogate for variation of substrate production and subsequent availability at coarse spatial and temporal scales (Reichstein et al., 2003; Davison et al., 2006) based on the strong positive correlations between canopy photosynthesis and LAI (Rodriguez et al., 2001; Reichstein et al., 2003; Barr et al., 2004; Ewert, 2004) and between photosynthesis and soil respiration (Janssens et al., 2001; Reichstein et al., 2003; Curiel yuste et al., 2004). For example, in a mixed temperate forest, a seasonal change in soil respiration was linearly correlated with a seasonal change in LAI (Curiel yuste et al., 2004). The linear correlation between soil respiration and LAI demonstrated in this study (Fig. 8c) supports use of LAI as an indicator of substrate availability.
Conclusions
Hurricane Frances caused defoliation and consequently reduced GPP and Re. The compensatory decline in both resulted in no significant change in NEP. The post-hurricane refoliation in the 2.5-month period following the hurricane compensated for only one fifth of the defoliation loss, though LAI recovered fully by the next growing season. GPP and Re also fully recovered within a year. Growth in elevated Ca did not alter the disturbance in damage and recovery. According to the findings from this study, hurricanes capable of causing significant defoliation with negligible damage to stems have negligible effects on NEP under current or future CO2-enriched environment.
Acknowledgements
This work was supported by a grant to the Smithsonian Institution by the US Department of Energy. We are grateful to Dr Ray Wheeler for his support. We appreciate comments from Drs John Erickson and DE Pataki and from two anonymous reviewers on an early version of this paper. We acknowledge the support of the Biomedical Office of NASA at the Kennedy Space Center, and the US Fish and Wildlife Service, Merritt Island National Wildlife Refuge.

