

Are Cape Ground Squirrels (Xerus inauris) Sensitive to Variation in the Pay-offs from Their Caches?
Abstract
For food caching to be adaptive, the benefits of recovery must outweigh the costs of storing an item. One of the costs to cachers is the risk of theft, and therefore, it is predicted that individuals may be sensitive to this theft and show various behavioural strategies to minimise it. In this study, we gave wild Cape ground squirrels (Xerus inauris) a choice between two different coloured items of the same food type: one item with a specific colour that was always artificially removed when cached and the other item with a different colour that was not removed when cached. During the choice presentations, subjects reduced the amount they consumed and cached of the food items with the colour that was experimentally removed when cached, despite the two items only differing in caching pay-off. This avoidance to choose the food with the colour that was removed occurred over time, which suggests that subjects were using information about the item's pay-off during cache recovery and this then impacted on successive decisions. This study highlights how the sensitivity to a food item's pay-off can affect an individual's choice towards items that offer the greater overall reward.
Introduction
Animals constantly face decisions that could have repercussions on their future fitness (McFarland 1977; Stevens 2008). One of the traditionally accepted assumptions of decision-making models is that evolutionary pressure has selected animals to behave ‘optimally’ (McFarland 1977; Kalenscher & van Wingerden 2011). An area that has been extensively studied is the optimal decisions animals make when foraging whereby animals are expected to maximise their rate of energy gain (Charnov 1976; Cowie 1977). In behaving optimally, individuals must balance the units of energy gain, such as intake rate against the cost of adopting that foraging strategy (Pyke et al. 1977; Pyke 1984). Costs can refer to a variety of factors such as predation risk (reviewed in Lima & Dill 1990), energetic pay-off (Richardson & Verbeek 1986) or competition (Lawlor & Maynard Smith 1976; Pimm & Rosenzweig 1981). The ability of an animal to forage optimally will therefore have a significant impact on their fitness (reviewed in, Pyke 1984).
One foraging strategy is to cache food upon discovery, storing it for later consumption (Vander Wall 1990) over both short and long periods (Cowie et al. 1981). Food caching in animals is only expected to occur where the pay-off to the cacher, in terms of recovering an item is greater than the investment in caching (Andersson & Krebs 1978; Stapanian & Smith 1984). Animals should therefore be sensitive to the pay-offs from their cached food items, altering their behaviour in relation to the net gains (Luo et al. 2014). Where cache loss occurs, species appear to engage in cache protection strategies to minimise this theft (Dally et al. 2006). Examples include being sensitive to the knowledge or attentiveness of audience members to a cache event (Bugnyar & Heinrich 2005; Samson & Manser 2015) or displaying deceptive strategies (Bugnyar & Kotrschal 2002; Steele et al. 2008). The type of cache protection strategy employed by a species is linked to a number of factors such as cognitive capabilities (e.g. scrub jays, Aphelocoma californica, Clayton et al. 2007), sociality (e.g. Ord's kangaroo rat, Dipodomys ordii, White & Geluso 2012) and cache type exhibited by that species (i.e. larder vs. scatter hoarding, Preston & Jacobs 2005; Zhang et al. 2011). One such cache protection strategy that is suggested to require advanced cognitive capabilities is exclusion performance.
Exclusion performance is defined as ‘selecting the correct alternative by logically excluding other potential alternatives’ (Call 2006; Mikolasch et al. 2012). One of the underlying assumptions of this exclusion performance ability is that animals must be spontaneous in their success (Call 2004), as gradual changes in decisions may suggest this behaviour is instead controlled by learning mechanisms (Schloegl et al. 2009). The ability to display exclusion in corvids has been linked to caching behaviour (Schloegl et al. 2009) where related non-caching species, such as jackdaws (Corvus glandarius), show an inability to successfully complete the same tasks (Schloegl 2011). In caching corvid species, exclusion may have evolved due to the cognitive demands associated with caching, whereby this exclusion behaviour allows individuals to keep track of the contents of their caches (Schloegl 2011; Mikolasch et al. 2012). However, this relationship has recently come under some scrutiny, where a study on another caching species, the Eurasian jay (Garrulus glandarius) did not find evidence of exclusion behaviour (Shaw et al. 2013). The inability of a species such as scrub jays to show spontaneous responses in cache tasks may be due to the fact that critical learning may occur at the time of recovery (Clayton et al. 2005). Therefore, animals may accumulate information about the profitability of a food item/site at recovery and use this information in subsequent cache decisions (Clayton et al. 2005).
We investigated whether manipulating the pay-offs of food items, within the context of food caching, would cause the Cape ground squirrel (Xerus inauris) to alter their caching behaviour. This species is an ideal study system to test this question due to their high degree of sociality (Waterman 1995), the harsh environment they live in (Tshikae et al. 2013) and their scatter-hoarding tendencies (Samson & Manser 2015). Together, these factors suggest that this species could be under intense competition for a limited amount of resources, which may select for sensitivity to conditions that lead to loss of these resources to competitors. Other rodent studies on food caching have highlighted a number of species to be sensitive to complete cache pilferage (where all items were removed, Huang et al. 2011; Luo et al. 2014), whereas here we tested whether subjects were sensitive to unequal cache pilferage rates between two items. We tested whether subjects would shift their behaviour in relation to the pay-off of the food item in order to choose items with the highest pay-off. Secondly, we predicted that this choice would be context specific, with subjects only showing variation in choice with respect to the food they cached and not consumed when food was provided. Huang et al. (2011) and Luo et al. (2014) suggested that the response of a rodent species to complete cache pilferage showed some degree of behavioural plasticity, but not at the level observed in some caching corvid species (Schloegl 2011). Therefore, thirdly, we predicted that our subjects would develop choice preference over presentation sessions, as individuals associatively learn the dichotomous cache pay-off of each item. Finally, we predicted that the variation in recovery potential of each of the two food colour types would cause individuals to show divergences in the spatial arrangements of caches in response to the perceived cache theft. It was shown in Merriam's kangaroo rats (Dipodomys merriami) that they switched from scatter to larder hoarding when cache theft was high, as concentrated stores were easier to defend (Preston & Jacobs 2001). However, Cape ground squirrels are highly social and have structured hierarchies (Samson & Manser 2015), meaning aggressive defence of single stores is unfeasible, due to the intensity of competition and the variation in competitive ability between individuals. Rather than larder hoarding the stolen food item type, we hypothesised that individuals would cache further away from their groups' central burrow and at lower densities to minimise cache loss (Clarkson et al. 1986).
Materials and Methods
Ethical Note
This work was undertaken in accordance with the guidelines outlined by the University of Pretoria, Animal Ethics Committee permit, no. ECO14-14.
Study Site and Species
This study was conducted at the Kuruman River Reserve, a research station located in the Kalahari Desert in South Africa (26°58′S, 21°49′E, Brotherton et al. 2001). At this site, Cape ground squirrels naturally occurred and a number of groups were habituated to close human observation (< 1 m) and all individuals within these groups had been given a unique dye mark (Samson & Manser 2015). This species is a highly social central place foraging sciurid that inhabits arid regions of southern Africa (Waterman 1995; Hillegass et al. 2008). Cape ground squirrels ‘scatter-hoard’ food around a central burrow and peaks in caching are observed after the heaviest periods of rainfall (J. Samson, unpubl. data). This species is also a short-term hoarder (Vander Wall 1990), as food caches are typically recovered food within 24 h of being made (Samson & Manser 2015). We tested 10 wild adult squirrels for this study (females = 6, males = 4) from six different social groups. Adult individuals were those over 12 mo of age, and due to large testis size, sex was easily determined (Waterman 1998). As the marked population has been maintained since May 2012, the age classes of all individuals used in this study were known (4 of 10 individuals where exact age was known, 6 of 10 individuals were present since the start of the project, being all older than 3 yr).
Experimental Design
In this study, we tested two experimental groups, ‘coloured removed’ and ‘non-coloured removed’, which corresponded to the food type that was removed after being cached by the animal during the manipulation phase (the other food item was not manipulated, Table 1). During the control condition, the observer manipulated no food items, but the coloured and non-coloured foods were presented. Each subject was assigned one of these two experimental groups using a randomised binary number generator, in a balanced design (n = 5 for each condition, Table 1). The food was standardised 2.5-cm-long monkey nuts (peanuts in the shell) that were either coloured or non-coloured. As no studies have been conducted on the visual abilities of this species, we assumed they are mono/dichromatic, as has been previously shown for other species of sciurids’ (Jacobs 1978). Therefore, in this study, we used blue dyed (the shell coloured with blue food dye) and uncoloured nuts, which would appear distinct from each other.
| Control | Manipulation | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| No caches removed | Caches of coloured or non-coloured food items removed according to experimental group. | |||||||||
| Presentation session | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| Coloured food removed | 3 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| 4 | 4 | 3 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | |
| 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | |
| 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | |
| 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | |
| Non-coloured food removed | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3 |
| 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | |
| 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | |
| 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | |
| 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | |
| Number of trials conducted | 39 | 40 | 39 | 40 | 40 | 40 | 40 | 40 | 40 | 39 |
| Total | 397 | |||||||||
- To control for experimental group effects, we had a balanced number of subjects (rows within the table) in both the non-coloured removed (n = 5) and the coloured removed (n = 5) groups. Within each presentation session, 3–4 trials were conducted.
Procedure
Subjects were presented with two identically sized food items, one coloured and one non-coloured nut, in a shallow cup attached to the end of a 1.5-m pole. Before the cup was placed on the ground, it was covered and shaken for 5 s to randomise the position of the two nuts. The cup was then placed on the ground, and the subject was allowed to approach the cup and choose an item, and once an item was chosen, the cup was lifted to stop the other item being taken. To reduce side bias, if the subject approached at an angle to the cup, it was lifted and placed again until the subject approached straight on (Fig. 1). This procedure was repeated up to four times for each subject during each presentation session (Table 1). When a food item was chosen, the choice (coloured or non-coloured) and fate (cached or eaten) were noted down. When subjects cached food items, a GPS point of the cache was taken. In addition, any caches (during the manipulation condition) of the removed item colour category were marked with non-descript cocktail sticks. As squirrels were never observed returning to caches during the period presentations were undertaken, individuals would therefore not associate the sticks with their caches. After the squirrels went below in to the sleeping burrow for the night, the cache sites for the removed items category were revisited and the food (and marker sticks) removed. When food was removed, the observer wore latex gloves to limit olfactory contamination of the cache site. All presentation sessions in this experiment were conducted in the afternoon, 1.5 h before individuals went below in to their burrows for the night to control for the motivational state of individuals. In addition, as subjects slept in the same burrow each night and returned to the burrow when they had ceased foraging, conducting presentations at this time allowed us to more easily locate the subjects. Sessions for each subject were terminated once four trials were completed or 30 min had passed since the start of the session. Trials were repeated within presentation sessions to ascertain average choice, rather than using a single trial. However, as caches were only manipulated after the presentation sessions had ceased, within these sessions no learning effects with respects to the item colour type's pay-off would occur.

Cache Fate and Placement Analysis
To determine whether caches were stolen by conspecifics or recovered by the cacher, we set up remote cameras near the site for 5 d and recorded 15-s videos whenever the infrared component was triggered (5210A series, LTL-Acorn Outdoors). As we could identify the individuals who removed caches, we could quantify an estimate for the proportion of caches recovered by the cacher or stolen by a conspecific. The distance caches were placed from the source (i.e. the centre of the burrow and where the choice task was presented) was determined by converting all coordinates from Cartesian to UTM and then calculating the Euclidean distance between the source and cache coordinates. Cache density was resolved by extracting within session nearest neighbour distances (NND) of caches for each subject animal. The NND was calculated using an in built function in the ‘spatstat’ package in R (Baddeley & Turner 2005).
Statistical Analysis
All statistical analyses were performed in R; release GUI, version 3.2.0 (R Core Team 2015). Generalised linear models (hereafter, GLM) with a Poisson error structure were used to analyse the number of food items chosen during the control condition. These models were conducted to determine whether there was any pre-existing preference for the caching or consuming the coloured or not coloured food items. Generalised linear mixed models (Bates et al. 2015) with a binomial error structure were used to analyse the choice data, as the outcome variable was binary (removed or non-removed items chosen). Condition (control or manipulation) and presentation session number (Table 1) were assigned as explanatory variables in the overall and presentation models, respectively. Experimental group (coloured or non-coloured food item removed), group and individual were assigned as random factors; in addition, trial number was included to control for repeated measures. As the number of individuals present in a group has been shown to negatively impact on the frequency at which food is cached as supposed to consumed (JS unpublished), we also included number of individuals as a random effect in the models. The number of individuals present refers to the total number of individuals counted by the observer at the group during that presentation session (all individuals were included in this count and not just those visible to the study subject). Linear mixed effect models were used to analyse the cache placement data, incorporating the same random effects as the first models. Significance of the explanatory variable for the mixed models was determined using likelihood ratio tests (hereafter, LRT, Crawley 2007), where models with the explanatory variables were compared to models with the explanatory variable removed. In all models, variance components were estimated using maximum likelihood methods, and additionally, all random effects were kept in the models. All p-values are two-tailed, and the significance level was set at 0.05.
Results
Across a total of 397 presentations, 230 food items were cached and 167 immediately consumed (number of items cached: total number of presentations, prop test; 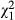 = 9.68, p = 0.002). Within the control condition, we found no pre-existing preference for caching either the coloured or non-coloured food items (GLM, z = 0.88, df = 19, p = 0.378) or a preference to consume either coloured food item (GLM, z = −1.21, df = 17, p = 0.227, Fig. 2). Subjects chose a higher proportion of items of the non-removed food colour type compared to the artificially removed food colour type comparing across conditions (proportion of non-removed/removed, manipulation: 177/141, control: 34/45, LRT,
= 9.68, p = 0.002). Within the control condition, we found no pre-existing preference for caching either the coloured or non-coloured food items (GLM, z = 0.88, df = 19, p = 0.378) or a preference to consume either coloured food item (GLM, z = −1.21, df = 17, p = 0.227, Fig. 2). Subjects chose a higher proportion of items of the non-removed food colour type compared to the artificially removed food colour type comparing across conditions (proportion of non-removed/removed, manipulation: 177/141, control: 34/45, LRT, 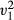 = 4.17, p = 0.041, Fig. 3a). No difference was observed in the proportion of food cached or consumed across conditions comparing both removed and non-removed items (LRT,
= 4.17, p = 0.041, Fig. 3a). No difference was observed in the proportion of food cached or consumed across conditions comparing both removed and non-removed items (LRT, 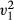 = 1.51, p = 0.219). Where animals chose the removed item colour type, no difference was observed in the proportion of food cached or consumed across conditions (LRT,
= 1.51, p = 0.219). Where animals chose the removed item colour type, no difference was observed in the proportion of food cached or consumed across conditions (LRT, 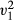 = 0.60, p = 0.438). Over manipulated presentation sessions, subjects chose a higher proportion of non-removed food colour type compared to the removed (LRT,
= 0.60, p = 0.438). Over manipulated presentation sessions, subjects chose a higher proportion of non-removed food colour type compared to the removed (LRT, 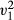 = 7.54, p = 0.006, Fig. 3b). No difference was observed in the proportion of food cached or consumed across manipulated presentation sessions comparing both removed and non-removed items (LRT,
= 7.54, p = 0.006, Fig. 3b). No difference was observed in the proportion of food cached or consumed across manipulated presentation sessions comparing both removed and non-removed items (LRT, 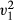 = 0.01, p = 0.920). In these manipulated presentation sessions, where animals chose removed items, no difference was observed between the number of items cached or consumed (LRT,
= 0.01, p = 0.920). In these manipulated presentation sessions, where animals chose removed items, no difference was observed between the number of items cached or consumed (LRT, 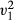 = 0.51, p = 0.474).
= 0.51, p = 0.474).


Comparing control to manipulated presentation sessions, items of both food colour types were cached at similar distances from the centre of their home range (Table 2). Over the manipulated presentation sessions, there was no change in how far food items of either colour category, the removed or non-removed items, were placed from the source (Table 2). We found a similar pattern with cache density, with no difference between the control and the manipulated presentation sessions in how caches were dispersed for either food colour type or whether density changed over trials (Table 2).
| Model | Estimate | SE |
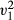
|
p |
|---|---|---|---|---|
| Distance from source | ||||
| Across treatments | 0.11 | 0.36 | 0.10 | 0.951 |
| Manipulated presentation sessions | 0.05 | 0.07 | 1.78 | 0.411 |
| NND | ||||
| Across treatments | 0.05 | 0.18 | −0.74 | 0.604 |
| Manipulated presentation sessions | 0.05 | 0.39 | 1.73 | 0.421 |
When quantifying the amount of caches recovered by the cachers or stolen by conspecifics, we found 27 of 47 caches (57.4%) were stolen and the rest were recovered by the cacher (proportion test; 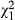 = 0.77, p = 0.382). Therefore, in this experiment, the two items have a predicted 43.6% (non-removed) to 0% (removed) return rate, accounting for this natural loss in food (a pay-off ratio of 0.44:0).
= 0.77, p = 0.382). Therefore, in this experiment, the two items have a predicted 43.6% (non-removed) to 0% (removed) return rate, accounting for this natural loss in food (a pay-off ratio of 0.44:0).
Discussion
This study demonstrates how Cape ground squirrels can alter their decisions regarding food item choice, developing a preference for the food item colour that offers the greater pay-off. As these pay-offs only differed in the context of caching and not consumption, subjects seemed to be developing a colour choice preference based on overall pay-off benefits. Over the course of the experimental condition, where caches were removed, the overall choice for the non-removed food items became stronger. Although subjects altered their food colour type choice, we observed no difference in the spatial arrangement of caches for the removed and non-removed food type.
Our results showed how by reducing the pay-off of a food colour type within a single context we could alter food choice behaviour in these ground squirrels. As the decision about what food colour type to choose did change over experimental presentations where caches were manipulated, we suggest that subjects were using the pay-off at cache recovery as a proxy for successive decisions with regard to colour choice. In other studies, there is still a debate as to the ability of animals to remember the content of their caches where a number of different types of food are cached (Clayton & Dickinson 1999a). We argue that the squirrels likely remember the contents of their caches and combine this with the information at recovery, that is whether a food item was there or not. The choice to avoid the removed food colour type over time might result from negative reinforcement that a specific colour offers no reward when cached. This reinforcement could arise when individuals return to an empty cache, triggering an inhibitory response through repeated negative associations (Pearce & Bouton 2001; Staddon & Cerutti 2003). This reduced pay-off when attempting to recover a removed item may cause an individual to develop some uncertainty about the overall pay-off of that food colour type (Stephens & Krebs 1986; Camerer & Weber 1992), which may then cause a shift in an individual's choice towards the food colour type that has a more certain pay-off, that is the most optimal choice (Caraco et al. 1980). The reason why we observed changes in choice in both contexts and not just caching is that subjects may assign a value to the reward and not the specific context in which the pay-off of these rewards differ (Sugrue et al. 2005). Similarly to other studies on rodents, despite the reduced item pay-off of one food type, subjects never ceased caching these unrewarded food colour types (Huang et al. 2011; Luo et al. 2014). One suggestion is that the motivational propensity to cache may be driven by an autonomous system irrespective of the costs vs. benefits of doing so (Clayton & Dickinson 1999b). Clayton & Dickinson (1999b) suggest that this compulsion to cache may be particularly prevalent when there are surplus resources or the animal is satiated, something which may be occurring in our study.
This preference for choosing the coloured food item with a positive association appeared to get stronger over the course of the manipulation condition. For exclusion abilities to be present, the shift towards picking the coloured food item type that offered the higher pay-off would have to occur after the first manipulation presentation sessions. Therefore, rather than subjects potentially showing such advanced abilities, the behavioural changes observed maybe mediated by associative learning processes (Shettleworth 1999). Although this remains to be tested, the ability of squirrels to learn such associations may suggest they are capable of using indirect information about a reward (Nawroth et al. 2014; Nawroth & von Borell 2015), in this case whether the colour was rewarded or not, and reuse such information in successive choice presentations. This will be of biological relevance to a caching species, as a single theft event (of an item) may not characterise the future theft potential of that item (Clayton et al. 2005).
Despite squirrels developing an avoidance of the removed coloured food type over time, we observed no shift in spatial arrangement of caches for when the removed or non-removed type of food items were cached. In Merriam's kangaroo rats, theft of caches by conspecifics causes individuals to shift their cache strategy from scatter to larder hoarding (Preston & Jacobs 2001). However, unlike Cape ground squirrels, Merriam's kangaroo rats are solitary living and can aggressively defend larder stores. Due to the Cape ground squirrels' sociality, aggressive defence of a larder is unfeasible, which may explain why we see no shift in the spatial arrangement of caches. Additionally, as we targeted all caches of the removed food colour type, there was no specific pattern of cache loss, such as the complete loss of food that was placed within a certain distance of the burrow system. Therefore, there are no specific stimuli for the squirrel's to learn in relation to the success of caches placed in different spatial arrangements.
In this study, we only compared two items, one that was not removed (but experienced normal rates of cache loss) and the other with complete artificial removal. This experimental design may make the choice decisions easier as there was a clear difference between the two items in terms of pay-off. Previous studies have highlighted how in two item choice tasks, subjects can weight choices against each other, showing a preference for the item with the greatest weighting component (Yechiam & Busemeyer 2006). In our example, as this study was undertaken in the wild, we could not control for natural rates of cache loss, and therefore, the two items did not have a 1:0 pay-off ratio. However, we were able to quantify an estimate for the cache loss and show that a preference does develop overtime given this 0.44:0 pay-off ratio. Further studies in the laboratory could determine how item preference develops with a varying pay-off ratio of the two food items, to pinpoint the threshold of response (Sugrue et al. 2005).
The economics behind the decisions of what to forage on are important for animals, particularly when these decisions can have fitness implications (Stephens & Krebs 1986; Kalenscher & van Wingerden 2011). We have shown how Cape ground squirrels are sensitive to the reward pay-offs of items, shifting decisions to choose the item with a higher pay-off. These decisions appeared to form through the squirrels' ability to associate colour with a reward pay-off. As the pay-off only differed in relation to caching and not consumption, it appears these squirrels are using a general association between a colour and a delayed reward to make subsequent choices, even though this pay-off only differed within the context of caching.
Acknowledgements
We would like to thank Tim Clutton-Brock and the Kalahari Research Trust for permission to work at the Kuruman River Reserve. Many thanks to the volunteers on the squirrel project for their help collecting the long-term data and Dave Gaynor for overseeing the project in the field. Research permits were obtained through Northern Cape Nature Conservation (permit no. FAUNA670/2013). We additionally thank Corina Logan and one anonymous reviewer for their helpful comments and suggestions. JS and MBM were both funded by the University of Zurich. The maintenance of the ground squirrel population is funded by the Universities of Cambridge and Zurich.
Conflict of Interest
The authors have no conflict of interest.

