

Resource-related variables drive individual variation in flowering phenology and mediate population-level flowering responses to climate in an asynchronously reproducing palm
Abstract
enMany tropical plant species show wide intra-population variation in reproductive timing, resulting in the protracted presence of flowering and fruiting individuals. Various eco-evolutionary drivers have been proposed as ultimate causes for asynchronous phenology, yet little is known about the proximate factors that control reproductive onset among individuals or that influence the proportion of trees producing new inflorescences within a population. We employed a nine-year phenological record from 178 individuals of the hyperdominant, asynchronously flowering canopy palm, Oenocarpus bataua (Arecaceae)¸ to assess whether resource-related variables influence individual- and population-level flowering phenology. Among individuals, access to sunlight increased rates of inflorescence production, while the presence of resource sinks related to current investment in reproduction—developing infructescences—reduced the probability of producing new inflorescences. At the population level, climate anomalies induced by El Niño Southern Oscillation (ENSO) affected the proportion of the population producing inflorescences through time. Moreover, the effects of ENSO anomalies on flowering patterns depended on the prevalence of developing infructescences in the population, with stronger effects in periods of low developing-infructescence frequency. Taken together, these results suggest that resource-related variables can drive phenological differences among individuals and mediate population-level responses to larger-scale variables, such as climate anomalies. Consequently, a greater focus on the role of resource levels as endogenous cues for reproduction might help explain the frequent aseasonal phenological patterns observed among tropical plants, particularly those showing high intra-population asynchrony.
Resumen
esMuchas especies de plantas tropicales exhiben bajos niveles de sincronía reproductiva dentro de sus poblaciones, y se caracterizan por la presencia constante de individuos en floración o fructificación. Varios fenómenos eco-evolutivos han sido propuestos como causas de estos patrones reproductivos, pero se desconocen los factores próximos que determinan el inicio de la floración en estas especies, o que influyen en la frecuencia de individuos iniciando su ciclo reproductivo a nivel de la población. Este estudio emplea un registro fenológico de nueve años, proveniente de 178 individuos de la palma de dosel, Oenocarpus bataua (Arecaceae)¸ para evaluar el rol del tamaño de la planta, el acceso a la luz, las anomalías climáticas, y los sumideros activos de recursos -en este caso, infrutescencias en desarrollo- como factores próximos de sus patrones fenológicos a nivel del individuo y de la población. Observamos efectos significativos entre variables relacionadas a la disponibilidad de recursos y los patrones de floración. A nivel del individuo, un mayor acceso a la luz incrementó la frecuencia de producción de inflorescencias, mientras que le presencia de infrutescencias en desarrollo disminuyó la probabilidad de producir nuevas inflorescencias. A nivel de la población, las anomalías climáticas incidieron en la frecuencia de individuos iniciando nuevas inflorescencias. Adicionalmente, observamos que la magnitud de los efectos climáticos en la frecuencia de individuos iniciando su ciclo reproductivo depende del porcentaje de la población con infrutescencias en desarrollo, con periodos de baja frecuencia de infrutescencias asociados a mayores incrementos. Enfatizamos que un mayor foco en el rol que los niveles de nutrientes a nivel del individuo juegan en la inducción del ciclo reproductivo puede mejorar significativamente nuestro entendimiento de patrones fenológicos no-estacionales en ecosistemas tropicales, como lo son los de floración asíncrona.
1 INTRODUCTION
The tropics harbor a great diversity of plant species that do not respond to seasonal climate and photoperiod as flowering cues. For example, some dominant tropical plant taxa exhibit mass flowering and fruiting patterns of irregular, supra-annual periodicity (i.e., masting) (Appanah, 1985, 1993). Other species reproduce several times a year at irregular time intervals that do not match seasonality in climate nor photoperiod (Bullock, Beach, & Bawa, 1983; Medway, 1972; Newstrom Frankie, & Baker, 1994a; Newstrom, Frankie, Baker, & Colwell, 1994b). Another common tropical phenological pattern is that of intra-population reproductive asynchrony, characterized by wide variation in reproductive rates and timing among individuals (Janzen, 1979). Asynchronously reproducing species are frequent in many communities (Hamman, 2004; Newstrom et al., 1994a, 1994b) and play important ecological roles within ecosystems, as the protracted presence of flowering and fruiting individuals can provide keystone plant resources to animals (Lambert & Marshall, 1991, Diaz-Martin, Swamy, Terborgh, Alvarez-Loayza, & Cornejo, 2014). While some tropical taxa use climatic conditions occurring sub- and inter-annually to cue reproduction in synchronous flowering events (e.g., Augspurger, 1983; Sakai et al., 2006; Wright, Calderón, & Muller-Landau, 2019), the proximate factors generating the wide variation in phenology among individuals of asynchronous species and the drivers of their population-level reproductive patterns remain unresolved. Identifying these proximate drivers would improve our ability to forecast temporal fluctuations in flower and fruit abundance in rain forest ecosystems and the aggregate responses of tropical plant communities to climate change.
Variation in resource acquisition and allocation may mediate reproductive asynchrony. Flower production and fruit production impose significant carbon and inorganic nutrient costs for most plants, requiring the allocation of current assimilates or depletion of stored reserves (Obeso, 2002; Reekie & Bazzaz, 1987; Stephenson, 1981). Some plants produce reproductive biomass in proportion to available resources (i.e., resource tracking, Fenner, 1998), but others may initiate flowering only after reaching threshold resource levels. Accordingly, some species halt flowering, sometimes for several seasons, following resource depletion in high seeding years (Crone, Miller, & Sala, 2009; Crone & Rapp, 2014; Miyazaki, Osawa, & Waguchi, 2009). Furthermore, molecular studies have demonstrated that resource levels play important roles as endogenous signals for reproductive development (Lastdrager, Hanson, & Smeekens, 2014; Ruan, 2014; Wahl et al., 2013). These observations suggest that individuals might initiate reproduction in response to endogenous factors signaling sufficient resources for flowering and fruiting (Isagi, Sugimura, Sumida, & Ito, 1997; Satake & Iwasa, 2000). If so, we would predict individual-level factors affecting resource acquisition, allocation and storage to cause variation in phenology.
Individual-level factors known to affect resource assimilation and allocation also influence plant phenological behavior (e.g., García León, Martínez Izquierdo, Mello, Powers, & Schnitzer, 2018; Martínez-Ramos, Anten, & Ackerly, 2009; Poorter et al., 2019; Visser et al., 2016). Among them, access to sunlight and size appear to play predominant roles in natural populations. Sunlight exposure increases photosynthetic rates, even in shade-tolerant species (Amadeu, Sampaio, & Santos, 2016; Chazdon & Pearcy, 1991), affecting the probability and timing of flowering, reproductive effort, crop sizes, fruit and flower abscission rates, and sex expression (Cai, 2011; Lorenzo et al., 2019; Poorter et al., 2019; Stephenson, 1981; Tucker-Lima, Caruso, Clugston, & Kainer, 2018). Additionally, resource allocation is often size-dependent (allometric), with larger plants directing a greater proportion of available resources to reproduction than smaller ones (Bonser & Aarssen, 2009; Tucker-Lima et al., 2018; Visser et al., 2016; Weiner, Campbell, Pino, & Echarte, 2009).
At the population level, researchers have emphasized the role of climate as a cue that synchronizes flowering among conspecifics (Mendoza, Peres, & Morellato, 2017; Rathcke & Lacey, 1985; Wright & van Schaik, 1994). However, climate also determines resource availability, either directly by mediating the supply of abiotic resources (e.g., water, sunlight) or indirectly through its influence on biotic processes that modify available resource pools (e.g., primary productivity, mycorrhizal function, and microbial activity) (Fernández-Martínez, Vicca, Janssens, Espelta, & Peñuelas, 2017; Högberg et al., 2010; Orwin et al., 2015; Richardson et al., 2005; Yang, Bastow, Spence, & Wright, 2008). In tropical rain forests, heavier rainfall at the end of the dry season may increase litterfall (Martínez-Yrízar & Sarukhán, 1990), greater cloud cover can decrease primary productivity (Graham, Mulkey, Kitajima, Phillips, & Wright, 2003), and higher temperatures can lead to greater soil nitrification and mineralization (Breuer, Kiese, & Butterbach-Bahl, 2002). Therefore, the climate can also influence plant phenology through its effect on resource budgets (Allen, Millard, & Richardson, 2017; Crone & Rapp, 2014; Satake & Iwasa, 2000).
Anomalies in El Niño Southern Oscillation (ENSO) generate dramatic examples of such associations. ENSO is an irregular fluctuation between warm and cold conditions in the tropical Pacific (El Niño/La Niña events respectively) that is responsible for the strongest patterns of inter-annual climate variation on Earth (McPhaden, Zebiak, & Glantz, 2006). El Niño events have been associated with forest-wide increases in net primary productivity (Asner, Townsend, & Braswell, 2000; Rifai et al., 2018), massive flowering and fruiting events (Chang-Yang, Sun, Tsai, Lu, & Hsieh, 2016; Chapman, Valenta, Bonnell, Brown, & Chapman, 2018; Wright & Calderón, 2006), and subsequent decreases in fruit production that can result in widespread frugivore famine under aggravating drought conditions (Wright, Carrasco, Calderón, & Paton, 1999). Consequently, through their effects on resource supply, ENSO-induced climate anomalies could affect the proportion of trees in a population having enough resources to initiate new inflorescences.
In this study, we use a 9-year phenological record from 178 individuals of the asynchronously reproducing palm Oenocarpus bataua (Arecaceae) to evaluate the influence of various factors on individual- and population-level flowering phenology. At the individual level, we (a) evaluate the effects of crown emergence through the forest canopy and tree height on inflorescence production among trees. If resource-related cues induced flowering, individual-level reproduction should be unconstrained or weakly constrained by the mild climate seasonality of the wet tropics within a year. Thus, we (b) evaluate whether individual-level inflorescence production tends to concentrate during certain months of the year. O. bataua can produce new inflorescences while bearing infructescences with developing fruits whose size, abundance, and high nutritional content upon ripening impose significant resource sinks on individual palms (Henderson, 1995). Therefore, we (c) evaluate whether the presence of developing fruits on individual trees affects the probability of producing new inflorescences. At the population level, we (d) characterize population-level phenology by computing the proportion of trees in different stages of their reproductive cycle (phenophases) throughout the study period. We then use this information to (e) assess whether ENSO-induced climate anomalies affect the proportion of individuals in the population initiating new inflorescences. Furthermore, if developing infructescences represent resource sinks that reduce the probability of inflorescence production at the individual level, their prevalence among individuals may affect the magnitude of population-level flowering responses to favorable climate anomalies. To examine this, we (f) assess whether the effects of ENSO anomalies on the proportion of trees initiating reproduction are mediated by the proportion of the population bearing developing fruits.
Previous analyses of O. bataua's phenology have described reproductive asynchrony among individuals and supra-annual peaks of flower and fruit at the population level (Núñez-Avellaneda & Rojas-Robles, 2008; Rojas-Robles & Stiles, 2009; Ruiz & Alencar, 2004). However, these studies have not simultaneously assessed individual-level variation in reproductive patterns and the link between population-level phenological behavior and inter-annual climate variability. Here, we resolve O. bataua's individual- and population-level phenological patterns to provide an assessment of the possible proximate factors influencing the phenological behavior of reproductively asynchronous tropical plants—an ecologically important, yet critically understudied, component of tropical forest communities.
2 METHODS
2.1 Study site
The study was conducted in a 130-ha study plot at the Bilsa Biological Station (hereafter BBS; 7900450W, 000220N; 436–615m elevation in our study site), a 3500-ha reserve of humid Chocó rain forest in northwest Ecuador (Figure S1). Average monthly temperatures at our study site vary between 26 and 28°C, while rainfall is markedly seasonal, with a 5-month dry season between July and November characterized by relatively low precipitation but persistent cloud cover.
2.2 Study species
Oenocarpus bataua is a hyperdominant, slow-growing, monoecious, protandrous Neotropical palm species reaching up to 35m in height (Henderson, 1995; ter Steege et al., 2013). O. bataua produces mass inflorescences over 2m long, with over 200 racemes bearing more than 300 flowers each. Following the appearance of an inflorescence bud, flowers take an average of 5 months to reach anthesis (Núñez-Avellaneda & Rojas-Robles, 2008; Rojas-Robles & Stiles, 2009). Individuals can bear multiple inflorescences simultaneously, and fertilized flowers develop large, lipid-rich, single-seeded drupes, comprising large crops of over 2000 fruits per individual. Fruits take approximately 13 months to develop following pollination (Rojas-Robles & Stiles, 2009). The abundance and nutritional content of its fruits and seeds make O. bataua's phenology an important determinant of food availability for large-bodied vertebrates, including humans (Henderson, 1995).
2.3 Phenological monitoring
We identified all adult O. bataua trees within our study plot (n = 178; Figure S1) and evaluated their reproductive status in monthly censuses (May 2008 to April 2017) where we counted the number of developing inflorescences, mature inflorescences (presence of open flowers), developing infructescences, and mature infructescences (presence of mature fruits) in each tree. We did not differentiate between male and female flowering phases. We added 64 individuals to our monitoring effort as we gradually expanded the study plot from 30ha to 130ha between January and September of 2010, and another 12 new individuals that reproduced for the first time through 2015.
2.4 Tree height and crown emergence
During a single census in 2013, we recorded the height of each Oenocarpus bataua tree in our study site by measuring the distance from its base to the tallest point of its crown using a laser range finder. Following the same procedure, the forest canopy height around each focal individual was measured as that of the tallest tree within a 10-m radius from the focal individual. We then assessed “crown emergence” for each focal tree by comparing its height with that of its surrounding canopy. A tree was categorized as “emergent” if taller than the canopy around it, and “non-emergent” otherwise. Because the presence of a taller tree within a 10-m radius does not necessarily guarantee that the focal tree will experience reduced access to sunlight, this approach has the limitation of underestimating the “true” number of focal trees with “emergent” crowns. However, this limitation only increases the probability of type II errors (“false negatives”), making our “emergence” metric a conservative estimate suitable for analysis.
2.5 ENSO and local climate
We evaluated whether ENSO plausibly drives climate variability in our site by correlating publicly available temperature, precipitation, humidity, and cloud cover data from TerraClimate (Abatzoglou, Dobrowski, Parks, & Hegewisch, 2018) and CRU TS v4.01 (Kobayashi et al., 2015), and the three ENSO indices most commonly employed to define El Niño/La Niña events: the Oceanic Niño Index (ONI), the Southern Oscillation Index (SOI), and the Multivariate ENSO Index (MEI) (https://www.esrl.noaa.gov/psd/enso, Smith, Reynolds, Peterson, & Lawrimore, 2008) (see Appendix S1 for a detailed description of the methods). Overall, El Niño conditions were significantly correlated with anomalously high temperature and vapor pressure levels in our study region, and moderate increases in rainfall and decreases in cloud cover (Table S1).
2.6 Individual-level analyses
 (1)
(1)The model included an interaction term of tree height and crown emergence (Height x Emergence), whose coefficient represents the difference in the effect of tree height for trees with emergent crowns compared to trees with non-emergent crowns. All predictors were centered at 0 and standardized (SD = 1) to aid comparison of effect sizes (Schielzeth, 2010) (summary statistics in Table S2). O. bataua's developing inflorescences are frequently depredated by insects, which can result in abortion (Pedersen & Balslev, 1992). In our study population, abortion rates averaged approximately 60 percent among individuals, and we estimate that an average of 35 percent of inflorescences per tree were attacked by insects (data not shown). However, if inflorescence abortion affected individual-level inflorescence production, we would expect abortion rates to be correlated with inflorescence initiation rates, yet we found no such association (r = −0.17, NS). Moreover, restricting the analysis to include only inflorescences that successfully flowered did not qualitatively affect our results.
 (2)
(2)We used production/no production of new inflorescence by focal individual i during month j as a binary response (n = 10,365), and the presence of developing infructescences in individual i in month j as a fixed effect factor. To test for temporal autocorrelation in the response, we computed partial autocorrelation correlation coefficients for 30 lags in the inflorescence production time series of all individuals in our sample, finding no significant patterns (see Figure S2 for details). Additionally, tree ID and year (indexed through individual i and year k, n = 122 and n = 10, respectively) were included as random intercepts, where
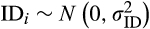 and
and
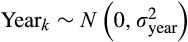 . Parameter estimates were generated using Laplace approximation, and the model was implemented using the “lme4” package version 1.1–21 in R (Bates, Mächler, Bolker, & Walker, 2014).
. Parameter estimates were generated using Laplace approximation, and the model was implemented using the “lme4” package version 1.1–21 in R (Bates, Mächler, Bolker, & Walker, 2014).
We assessed seasonal constraints in reproduction by testing for clustering of reproduction within a year at the individual level. To do this, we computed 95% confidence intervals of the expected number of months in which a given number of reproductive events would occur if flowering onset was equally likely each month of the year (see Figure S3 for details and a schematic description of the process). We complemented this analysis by evaluating how the probability of inflorescence production among individuals (n = 153) varied by month of the year (January–December) using a GLMM with production/no production of inflorescences each month as a response, month of the year as a fixed effect factor, and tree ID as a random effect (see Table S3 for details).
2.7 Population-level analyses
We calculated the proportion of trees in each phenophase every month between May 2008 and April 2017. Prior to analyzing the effects of ENSO anomalies on population-level flowering phenology, we temporally downscaled our data from monthly observations (n = 108) to non-overlapping trimesters (4-month period averages, n = 27) for two reasons. First, the monthly proportion of trees bearing developing inflorescences showed a complex autocorrelation structure with significant lags of up to 20 months. Decreasing the temporal resolution of our data yielded significant partial autocorrelation coefficients only for the first two lags. Second, monthly variation in ENSO indices showed weaker correlations with climate than did 4-month periods (Table S1). Because the proportion of trees producing new inflorescences was strongly autocorrelated between consecutive months (r = .92), we imputed missing observations in December 2008, October and November 2012, December 2014, and April 2015, by assuming a linear rate of change between two months of observation bounding a missing value. We then computed the non-overlapping trimestral time series using the full, imputed monthly time series.
 (3)
(3)We used the proportion of trees bearing developing inflorescences in the focal trimester as a response (Inflort) and included the value of the response for the previous trimester as a predictor to account for temporal autocorrelation (Inflort-1). We selected the ENSO index most strongly associated with climate in our site (MEI) and selected between the two time lags of the MEI most correlated with local climate (lag 0: MEIt, lag 1: MEIt-1) by running two models, including only one of them as a predictor, and selecting the model with the lowest AIC score (MEIt). We used ENSO indices instead of the climate variables from TerraClimate because ENSO simultaneously affects multiple climate variables, providing an integrated “climate package” (Stenseth et al., 2003). To evaluate whether the prevalence of individual-level resource sinks affects population-level flowering phenology, the model included the proportion of trees bearing developing fruits in the population (GFt) as a predictor. We also included an interaction term between MEIt and GFt, which models the linear dependence of the phenological effects of MEIt on GFt or (alternatively) the dependence of GFt effects on MEIt. We visualized this interaction by computing a cross-sectional plot of the effect of MEIt on the response at three reference (standardized) values of GFt (−1, 0, 1) using the “visreg” package in R (Breheny & Burchett, 2013). To do so, each standardized observation of GFt (a continuous variable) was assigned to the reference value closest to it. We standardized all predictors on Eqn. 3 to a mean of 0 and standard deviation of 1 except for MEI, which is a standardized index (summary statistics in Table S2). We detected no significant autocorrelation in model residuals (Box–Pierce test: p > .05 for all lags; Godfrey, 1979).
All data manipulation, visualization, and analysis for this study were carried out using R version 3.6.1 (R Core Development Team, 2018).
3 RESULTS
3.1 Individual-level predictors of phenological variation
Our model (Equation 1) explained a significant proportion of the variation in reproductive rates in our sample (R2 = .22, df = 149, F = 13.67, p < .001). For average tree heights, canopy emergence was estimated to increase yearly inflorescence production by 0.32 (i.e., 34.7 percent) compared with trees whose crowns were under the canopy (β1 = 0.32, SE = 0.08, p < .001; Figure 1a). Inflorescence initiation showed a significant positive relationship with tree height for trees with non-emergent crowns, with an average increase of 3.5m in tree height (equal to 1SD) predicted to increase yearly inflorescence production by 0.21 inflorescences per year (β2 = 0.21, SE = 0.07, p = .002; Figure 1b). In contrast, we detected a significant negative interaction of similar magnitude between tree height and crown emergence (β3 = −0.23, SE = 0.08, p = .006), indicating that tree height was only associated with higher reproductive rates for trees with canopy-covered crowns (Figure 1b). The presence of developing infructescences significantly reduced the probability that an individual would produce new inflorescences each month from 0.117 to 0.094 (i.e., a 20.4 percent decrease) (exp(β1) = 0.796; Table 1).

| Random effects | ||
|---|---|---|
| Number of blocks | SD | |
| ID | 122 | 0.255 |
| Year | 10 | 0.310 |
| Fixed effects | ||||
|---|---|---|---|---|
| Coefficient | SE | Z | p | |
| Intercept | −2.14 (0.117) | 0.110 | −19.50 | <.001 |
| GF present | −0.228 (0.796) | 0.073 | −3.12 | .002 |
| AIC | Log likelihood | Deviance | DF residuals | |
| 6,999.6 | −3495.8 | 6,991.6 | 10,361 | |
The number of distinct months of reproductive onset did not differ from the random expectation for 151 of 153 trees (98.7 percent of the sample; Figure 2), suggesting that individuals do not consistently initiate flowering in certain months of the year. Accordingly, we found no significant differences in probability of inflorescence production among trees for 10 out of 11 months relative to the reference month of July (Table S3). Though the estimated probability that an individual would produce a new inflorescence in December was 48.9 percent lower than in July (0.103 versus. 0.053; Table S3), the lack of significant differences for all other months points to an aseasonal pattern of inflorescence production.

3.2 Population-level phenological behavior
Our sample captured three population-level reproductive cycles, with supra-annual reproductive peaks separated by periods of approximately 30 to 36 months. The proportion of trees bearing mature fruits (mean ± SD =5.1 ± 6.7 percent, range = 0 – 27.0 percent) surged around 18 to 24 months following peaks in inflorescence initiation, with individuals bearing mature fruits present throughout the majority of the study (85 percent of the monitoring period; Figure 3). The proportion of individuals bearing developing inflorescences remained consistently above 13 percent of the population, and well over 20 percent for most of the study period (mean = 36 percent; Figure 3, Table S2).

3.3 ENSO anomalies and population-level phenology
Our model (Equation 3) explained most of the variation in the proportion of trees initiating new inflorescences through time (R2 = .86, df = 21, F = 32.75, p < .001). Under average ENSO conditions, developing inflorescence frequency was negatively correlated with developing-infructescence frequency (Figure 4a; β3 = −0.052, SE = 0.012, p < .001), with increases of 12 percent in the proportion of trees with developing fruits (equal to 1 SD) associated with a 5.2 percent decrease in the proportion of trees bearing developing inflorescences. In contrast, we detected a positive relationship with the Multivariate ENSO Index (MEI) for average levels of developing-infructescence frequency (Figure 4b; β2 = 0.056, SE = 0.013, p < .001), indicating a link between the warm phase of ENSO, typically associated with El Niño conditions, and a greater frequency of trees with developing inflorescences. The effects of the MEI on the frequency of trees initiating reproduction depended significantly on the frequency of trees bearing green fruits (Figure 4c; β4 = −0.040, SE = 0.011, p = .001). During periods of low green fruit frequency (1SD below average, ~13 percent), the model predicted that a change from −1 to 1 in the MEI (moderate la Niña to moderate el Niño conditions) would increase the proportion of trees bearing developing inflorescences by 19 percent, compared to a change of only 3 percent in periods of high developing-infructescence frequency (1SD above average, ~37 percent).

4 DISCUSSION
Asynchronously flowering species are common in the tropics, with estimates as high as 22 percent of all species sampled in some tree communities (Hamann, 2004). While many eco-evolutionary drivers have been proposed as ultimate causes for asynchronous phenological patterns (Augspurger, 1981; Elzinga et al., 2007; Janzen, 1971), the proximate factors controlling individual- and population-level patterns are poorly understood. By studying the asynchronous canopy palm Oenocarpus bataua, we found that access to sunlight and resource sinks in the form of developing infructescences affect patterns of inflorescence production among individual trees, and interact with ENSO-driven climate variation to shape the flowering phenology of the population as a whole. Our results provide one of the first assessments of the factors controlling flowering phenology across individuals and populations of asynchronous species and suggest that a resource-centric view of phenological variation might help elucidate the asynchronous phenological patterns frequently observed in tropical plant species.
4.1 Individual-level patterns
Reproductive asynchrony could emerge if individual-level differences (in environmental stress, access to resources, genetics, etc.) affect the rate of exposure or the sensitivity of individual plants to relevant environmental cues (Ollerton & Lack, 1992). However, this mechanism predicts variation in flowering time only within seasons in which threshold levels of a cue occur. For example, species flowering in response to spring warming may vary in flowering time only within the spring, while those cued by threshold levels of solar irradiance might vary in flowering time only within seasons of sufficient day length. As a result, variation in exposure to external cues fails to explain the degree of reproductive asynchrony observed in many tropical species. In O. bataua, individuals in proximity and under similar microsite conditions routinely initiate flowering out of phase with one another. Moreover, we found no evidence of seasonality in flowering onset among individuals in our focal population. Together, these observations show that external cues are unlikely drivers of reproduction in O. bataua.
Instead, individual-level differences can influence reproduction through their effect on resource budgets. Access to sunlight influences photosynthetic rates and carbohydrate production and enhances various components of plant reproduction, including reproductive frequency and crop sizes (Poorter et al., 2019). Therefore, reproductive rates may be higher among individuals with greater access to sunlight. Accordingly, we found that crown emergence from the canopy was associated with a 34.7 percent increase in inflorescence production controlling for differences in plant size, suggesting that photosynthetic rates may drive phenological variation among individuals in this species. Additionally, tree height only had significant effects among individuals with non-emergent crowns (Figure 1a). These results suggest that tree height might influence rates of reproduction through increased access to sunlight rather than through allometric effects on resource allocation in O. bataua, though it is possible that size-dependent resource allocation could affect other reproductive variables, such as crop sizes or fruit abortion rates. Although our methodology establishes a link between sunlight and phenological behavior in O. bataua, we used a conservative method to estimate crown emergence. Direct measurements of the amount of solar radiation in treecrowns would likely reveal even greater effects of access to sunlight on inflorescence production rates.
If resource levels cue flowering among individuals, active resource sinks could reduce the probability that a tree will develop new inflorescences (Obeso, 2002). Accordingly, the presence of developing infructescences in an individual led to an estimated reduction of 20.4 percent in the monthly probability of inflorescence production. Moreover, increases in the proportion of trees bearing developing infructescences suppressed the population-level increases in inflorescence production associated with the warm phase of ENSO (below), suggesting that developing infructescences act as resource sinks that influence individual-level phenological behavior. Because the presence of developing fruit is a direct consequence of previous inflorescence production, it is possible that the decrease in the probability of producing new inflorescences reflects resource expenditures associated with recent inflorescence production. However, the significant increase in reproductive biomass associated with the transition of flowering to fruiting, as well as the high nutritional content and long maturation time of its fruits, suggests that infructescence development likely demands a greater amount of resources than inflorescence development, making fruit production and ripening stronger resource sinks than inflorescence production.
Our results support the hypothesis that resource status might mediate phenological behavior in O. bataua by providing endogenous flowering cues (Lastdrager et al., 2014; Ruan, 2014; Wahl et al., 2013). However, further analyses, such as experimental manipulation of inorganic resources, direct measurements of non-structural carbohydrates, nitrogen, and phosphorous concentrations before and after reproduction (e.g., Miyazaki et al., 2009), or developing-fruit pruning treatments, are necessary to conclusively establish the role that resource levels and resource sinks play on reproductive onset in our system.
4.2 Population-level patterns
O. bataua is pollinated by a diverse array of insect species (Núñez-Avellaneda & Rojas-Robles, 2008), and many vertebrate taxa consume its fruits and lipid-rich seeds (Henderson, 1995). We observed a constant presence of flowering individuals and a protracted presence (85% of the study period) of trees with mature fruit. Consequently, although the presence of fruiting trees was not constant, we propose that O. bataua's hyperdominance (ter Steege et al., 2013), massive flower and fruit crops, protracted presence of flowering and fruiting trees (with pronounced peaks lasting ~1.5 years), and the wide array of frugivores and pollinators it sustains (Mahoney et al., 2018; Narasimhan, unpublished Data), makes it a keystone plant resource for many rain forest organisms of the Chocó bioregion (Terborgh, 1986, Diaz-Martin et al., 2014).
While reproduction had no apparent seasonality at the individual level, the frequency of trees producing new inflorescences in the population showed supra-annual peaks occurring at 20- to 36-month intervals (Figure 3). Although some masting tropical taxa use supra-annual climate anomalies to cue reproduction in mass fruiting and flowering events, they are characterized by high levels of synchrony among individuals (Sakai et al., 2006). In contrast, the proportion of reproducing trees in our focal population never dropped below 12 percent, with about a third of the population, on average, bearing developing inflorescences during any given month. Therefore, it is unlikely that the supra-annual phenological patterns reported in this study stem from supra-annual climatic cues.
In contrast, climate variation can affect reproductive patterns through its effects on resource supply (Allen et al., 2017; Pau et al., 2013; Wright & Calderón, 2006). We found that the proportion of trees producing new inflorescences was influenced by ENSO, with anomalously warm conditions (El Niño) associated with higher inflorescence initiation in our study population. Additionally, the proportion of trees with developing fruits was negatively associated with that of trees initiating new inflorescences, with higher developing-infructescence frequencies suppressing the flowering response of the population to ENSO-induced climate anomalies. These results show that inter-annual climate variation can influence temporal patterns of reproduction at the population-level in asynchronous species and that the magnitude of these effects is mediated by the prevalence of individual-level resource sinks in the population—in this case, developing infructescences. Though ENSO anomalies seemed significantly associated with changes in temperature, rainfall, and cloud cover in our study site (Table S1), we did not directly measure local climate variables and cannot currently discern the precise climatic effects of ENSO anomalies nor the variables responsible for O. bataua's population-level responses. Therefore, direct measurements of local climate, coupled with experimental warming (e.g., Nakamura, Muller, Tayanagi, Nakaji, & Hiura, 2010), irrigation (e.g., Wright & Calderón, 2006), or high-intensity light treatments (e.g., Graham et al., 2003), could help identify the climate variables that generate population-level reproductive responses in O. bataua.
Although O. bataua's population-level phenology responded to inter-annual variation in climate, other species are likely to respond to climate fluctuations occurring at different temporal scales. O. bataua produces resource-demanding reproductive structures that develop over multiple years (Rojas-Robles & Stiles, 2009), and shorter-lived climate fluctuations, such as those occurring seasonally, are unlikely to affect resource budgets enough to induce detectable responses in the population. In contrast, many asynchronous species produce flower and fruit crops with lower energetic demands that can develop within a single season (e.g., Milton, Windsor, Morrison, & Estribi, 1982), or multiple times per year (e.g., Wright & Calderón, 2018). Consequently, climate fluctuations occurring over shorter temporal scales might provide resource pulses of sufficient magnitude to influence the frequency of individuals initiating reproduction. This mechanism could explain why the populations of many species show annual reproductive peaks even though the timing of reproduction within the year varies widely among individuals (e.g., Wright et al., 2019).
5 CONCLUSIONS
In asynchronously reproducing species, the protracted presence of flowering individuals suggests that climatic and photoperiodic cues are unlikely to control flowering onset. In contrast, the role of resource levels as flowering cues predicts that factors related to resource assimilation and allocation should generate phenological variation among individuals. Our study shows how differences in access to sunlight result in phenological variation among individuals of an asynchronously reproducing canopy palm. Additionally, we show that ENSO-induced climate anomalies, factors likely affecting resource supply for the whole population, influence population-level flowering phenology and that the magnitude of their effect is mediated by the prevalence of resource sinks among individuals. Further studies are needed to conclusively demonstrate that resource levels induce flowering and the mechanisms by which climate anomalies affect resource supply and phenology in O. bataua. Nevertheless, our results provide one of the first assessments of the proximate drivers of phenological patterns for individuals and populations of an asynchronous species, underscoring how resource dynamics across levels of organization might drive the weak reproductive synchrony among conspecifics observed in many tropical plant taxa.
ACKNOWLEDGEMENTS
We are grateful for the support of FCAT (Fundación para la Conservación de los Andes Tropicales), the Jatun Sacha Foundation, and the Ecuadorian Ministry of the Environment. We thank J. Olivo, F. Castillo, M. Gonzalez, N. Gonzalez, M. Baquero, and N. Oleas for logistical assistance; E. Iverson, the Karubian Lab, and the Mazer Lab for comments on earlier drafts of this manuscript; and two anonymous reviewers for their thoughtful feedback. This project was supported by: The Conservation, Food & Health Foundation; Disney Conservation Fund; National Science Foundation (EAGER #1548548 and a Graduate Research Fellowship to LMB: DDIG #1501514); National Geographic Society; Tulane University; and the United States Fish & Wildlife Service (NMBCA # 6318). All research was conducted with approval of the Ecuadorian Ministry of the Environment (MAE-DNB-CM-2015-0017).
Open Research
DATA AVAILABILITY STATEMENT
Data are available from the Dryad Digital Repository: https://doi.org/10.25349/D95P5W (Ramirez-Parada, Cabrera, Diaz-Martin, Browne, & Karubian, 2020).

