Department of Chemistry, Alberta Ingenuity Centre for Carbohydrate Science, University of Alberta, Edmonton, Alberta, Canada T6G 2G2
Christopher W. Cairo, Department of Chemistry, Alberta Ingenuity Centre for Carbohydrate Science, University of Alberta, Edmonton, Alberta, Canada T6G 2G2
David E. Golan, Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, 250 Longwood Avenue, Boston, MA 02115
Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, 250 Longwood Avenue, Boston, MA 02115
Hematology Division, Brigham and Women's Hospital, Boston, MA
Christopher W. Cairo, Department of Chemistry, Alberta Ingenuity Centre for Carbohydrate Science, University of Alberta, Edmonton, Alberta, Canada T6G 2G2
David E. Golan, Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, 250 Longwood Avenue, Boston, MA 02115
Department of Chemistry, Alberta Ingenuity Centre for Carbohydrate Science, University of Alberta, Edmonton, Alberta, Canada T6G 2G2
Christopher W. Cairo, Department of Chemistry, Alberta Ingenuity Centre for Carbohydrate Science, University of Alberta, Edmonton, Alberta, Canada T6G 2G2
David E. Golan, Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, 250 Longwood Avenue, Boston, MA 02115
Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, 250 Longwood Avenue, Boston, MA 02115
Hematology Division, Brigham and Women's Hospital, Boston, MA
Christopher W. Cairo, Department of Chemistry, Alberta Ingenuity Centre for Carbohydrate Science, University of Alberta, Edmonton, Alberta, Canada T6G 2G2
David E. Golan, Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, 250 Longwood Avenue, Boston, MA 02115
This article was originally published online as an accepted preprint. The “Published Online” date corresponds to the preprint version. You can request a copy of the preprint by emailing the Biopolymers editorial office at [email protected]
INTRODUCTION
Cell surface adhesion receptors have two major functions. First, these molecules positively and negatively regulate the binding of a cell to other cells or to solid-phase supports (e.g., extracellular matrix), controlling the formation and dissolution of both stable (e.g., tight junction) and dynamic (e.g., focal adhesion) adhesive structures. Second, these receptors are critical signaling molecules in the plasma membrane, facilitating the exchange of information between cells (e.g., immune synapse). Unlike receptors that interact with freely diffusing small molecules, adhesion receptors typically function by interacting with a counter-receptor or ligand on an opposing cell or solid-phase support. Because both binding partners are generally confined to the plasma membrane, these interactions are acutely sensitive to the peculiarities of the membrane environment. The restriction of receptors to the two-dimensional, viscous, and heterogeneous plasma membrane slows their diffusion and therefore the rate of the biological events they mediate.1 Measurements of receptor diffusion in the plasma membrane have consistently shown that these molecules experience dramatically slowed diffusion, even when the viscosity of the environment is taken into account.2 A variety of mechanisms have been found to contribute to this phenomenon of anomalous diffusion within the plasma membrane (vide infra), and these mechanisms provide a means of specifically regulating the lateral mobility of cell surface molecules. As a central parameter in determining the rate of biological interactions, the two-dimensional diffusion, or lateral mobility, of these receptors is a critical determinant of their role in cellular biology.
Biological Function of Lateral Mobility
The range of two-dimensional diffusion coefficients that have been measured for membrane proteins and lipids spans several orders of magnitude. Lipids generally diffuse relatively rapidly in the plasma membrane, with diffusion coefficients close to 1 × 10−8 cm2 s−1 (1 μm2 s−1), whereas transmembrane adhesion proteins generally diffuse relatively slowly, with diffusion coefficients as low as 1 × 10−11 cm2 s−1 (0.001 μm2 s−1).3, 4 But how do these values affect receptor function? A useful starting point is to consider the surface area coverage of a freely diffusing molecule in the plasma membrane. We approximate the surface area, SA, of a simple spherical cell, such as a human T cell (∼3 μm in radius, r), as
()
We further assume that the area of coverage, A, for a particle diffusing in two dimensions with a two-dimensional diffusion coefficient D is given by
()
Rearrangement of Eqs. (1) and (2) allows estimation of the time required for a particle experiencing free diffusion to explore an area equivalent to the cell's surface area
()
The measured diffusion coefficients of membrane proteins and lipids span three orders of magnitude, and the time required for a molecule to explore the surface area of a cell would also be expected to vary over this range—for our model cell, the range is from 2.8 × 101 to 2.8 × 104 s (or about 30 s to 8 h). Similar considerations are expected to govern the rates of collision of two laterally mobile molecules on the cell surface.5
This oversimplified model of protein and lipid diffusion in the membrane provides a sense of the timescale over which these interactions take place. However, it is important to note that molecules in the membrane often experience anomalous diffusion because of complex mechanisms which are still not completely understood.3 A number of important molecular and cellular events require the lateral motion of membrane proteins and lipids, and therefore depend on receptor lateral mobility. These phenomena include receptor clustering, microdomain formation, cell adhesion, and receptor-ligand interactions among others.4, 6, 7 Therefore, accurate measurements of lateral mobility are required to understand these events at the molecular level.
Methods for Studying Lateral Mobility
Investigation of cell surface receptor lateral mobility requires specific labeling of the molecules using fluorescent dyes or other optical labels (e.g., nanoparticles). The primary methods for determining receptor lateral mobility are fluorescence microscopy (FM), fluorescence photobleaching recovery (FPR; also known as fluorescence recovery after photobleaching, FRAP), and single-particle tracking (SPT). Each method has particular advantages, and the three approaches are in many ways complementary to one another.
FM allows for whole-cell and subcellular imaging of the localization of surface molecules. This information can be used to obtain an estimated or qualitative measure of receptor lateral mobility.8, 9 Quantitative methods for analyzing FM mobility data are a more recent application, and these methods offer insight into the diffusion coefficient of receptors in many regions of the cell simultaneously.10, 11 Fluorescence correlation spectroscopy can also be used to achieve quantitative diffusion measurements.12
FPR is an established quantitative method for measuring membrane protein and lipid diffusion. Early work with this method provided crucial insight into the physical properties of membrane proteins.13, 14 Observations have consistently shown that proteins in biological membranes have low lateral mobility relative to proteins and lipids in model membranes. Additionally, experiments on many cell surface proteins indicate the presence of an “immobile fraction”—a population of receptors that have diffusion rates substantially slowerthan those of a rapidly diffusing, mobile population of molecules.
Both FM and FPR determine the average lateral mobility of an ensemble of molecules by observing a large population of receptors. Nonensemble and single-molecule methods have significant advantages in measuring receptor mobility. SPT has become an important complementary method for the analysis of receptor diffusion.3 SPT is unique in providing diffusion measurements of single molecules, and therefore can provide insight into the properties of poorly represented molecular populations or rare molecular events. Additionally, compared with FPR, SPT has increased sensitivity for measuring low diffusion coefficients.15, 16 SPT methods can also be integrated with laser optical tweezers systems to allow for micromanipulation and force measurements.17
Factors Contributing to Lateral Mobility
Physical factors that affect membrane protein lateral mobility include the size (cross-sectional area) of the protein, the viscosity and other properties of the membrane,2, 18 interactions of the protein with other membrane-associated proteins,19, 20 and the total concentration of proteins in the membrane (i.e., molecular crowding).21
Some of these factors, such as the size of a protein, would be expected to remain constant for a transmembrane receptor. Other factors may change rapidly on the biological time scale. The membrane environment in which a protein resides could change, perhaps as the protein enters a microdomain22 or when the cell radically alters its membrane composition.23 Changes in receptor surface expression are also known to occur rapidly in response to signaling.24 However, the most dramatic alterations in lateral mobility are typically due to changes in molecular associations. In particular, large changes in receptor mobility often occur when the receptor alters its state of association with the cytoskeleton—a network of crosslinked and generally immobile proteins at the inner surface of the plasma membrane—or with extracellular ligands such as those in the extracellular matrix (ECM).20, 25
Binding to Cytoskeleton or Associated Proteins
Many transmembrane proteins interact with the cytoskeleton via their cytoplasmic domains.26, 27 In systems where this occurs, the lateral diffusion of the protein may be reduced by more than an order of magnitude. Therefore, changes in receptor interactions with cytoskeleton-associated proteins can be a determining factor in receptor lateral mobility. A variety of cytoskeleton-associated proteins have been shown to alter transmembrane protein lateral diffusion through specific binding interactions.
A well-known example of a cytoplasmic binding domain that affects receptor lateral mobility comes from work on MHC proteins with engineered cytoplasmic tail mutations. Although some systems show little change upon cytoplasmic tail deletion,28 others show dramatic changes in receptor lateral mobility depending on the specific sequence29 and length of the cytoplasmic domain.28, 30 This general mechanism influences the mobility of a large variety of immune cell receptors required for T cell activation.31-33
Although cytoplasmic domain binding sites are clearly important for receptor interactions with cytoskeleton-associated proteins, it should be noted that these sites are often not static entities. Chemical and conformational changes can influence the binding interaction.34-37 For example, measurements of the two-dimensional affinity of receptors within a contact zone38 not only show deviations from the three-dimensional (solution-phase) interaction of the same receptors,1, 39, 40 but also are consistent with conformational changes in the activated receptor leading to increased receptor affinity and increased interaction with cytoskeletal regulator proteins.41
Receptor Clustering
The clustering of cell surface receptors is known to effect signaling,7 but what is its effect on receptor lateral mobility? Receptor complexes, or clusters, would be expected to have reduced lateral mobility due to an increase in cross-sectional area. The Saffman and Delbrück model predicts only a weak dependence of lateral mobility on cross-sectional radius, a,
,2 although more recent treatments suggest a stronger dependence .18 From both models, we would predict that large receptor clusters would exhibit reduced lateral mobility. The formation of small clusters (e.g., dimers or trimers) might be difficult to discern in a live cell, however. In general, there are not many examples of large changes in receptor mobility due to clustering alone,42 and model studies of receptor aggregation using single-particle tracking have shown only small changes in lateral mobility.43
Whereas cluster formation may not substantially alter lateral mobility, changes in lateral mobility can significantly restrict the ability of a receptor to form clusters. As noted earlier, cytoplasmic or extracellular contacts can restrict the mobility of transmembrane receptors. The maintenance of these contacts can prevent receptors from clustering, and their release can promote cluster formation. Therefore, knowledge of the lateral mobility of a receptor and the mechanism of its regulation can provide insight into events that require homo- or hetero-clustering of the receptor within the plane of the membrane.44
Microdomains
The lateral mobility of membrane components is highly dependent on membrane viscosity. Most models predict an inverse relationship between D and the viscosity of the membrane, μ,
.2, 18 The situation is complicated, however, by the potential for heterogeneity within the membrane in the form of lipid rafts6 or other microdomains.16, 45 Work in model systems has clearly established the inherent preference of certain lipids to associate with one another,46 although direct observation of this phenomenon in vivo has been challenging.6 Membrane components diffusing through such microdomains could exhibit altered lateral mobility if the viscosity of the domain is significantly different from that of the surrounding lipid. Observations suggestive of this phenomenon have been found using single-molecule methods in live cells.22
T Cell Surface Receptors
As noted earlier, a large number of physical factors regulate the lateral mobility of transmembrane proteins. Lateral mobility is a central parameter in determining the dynamics of adhesion systems. The human T cell presents an exceptionally well studied model for examining the role of this parameter in a diverse array of cellular interactions. Because the T cell takes part in many different and varied adhesive interactions, we shall not describe T cell adhesion comprehensively but instead focus on several examples in which molecular regulation of lymphocyte receptor lateral mobility has been shown to play a role in receptor function. Although our exemplary systems have been well described in previous reviews of T cell function (see reviews31-33, 47, 48), we aim here to propose a general framework for classifying lateral mobility mechanisms that regulate T cell adhesion systems. We define four primary classes of mechanisms for regulating the lateral mobility of receptors in the T cell plasma membrane: reorganization, recruitment, dispersion, and anchoring.
REORGANIZATION
The lateral association of receptors with specific lipids, glycans, or other proteins can alter receptor conformation or lead to tertiary interactions, both of which can affect receptor function. Membrane components can reorganize laterally through the formation of lipid rafts and other microdomains and through homo- and hetero-clustering events.
Reorganization: Change in the specific lateral associations of a receptor with other membrane components, including both lipids and proteins. (Figure 1)
1. Lateral reorganization of membrane components. A: Dissociated, laterally mobile membrane receptors (blue) and co-receptors (purple). B: Membrane receptors can diffuse laterally to form homo- or hetero-clusters, or can associate with lipid microdomains (orange).
Probably the best known example of lateral organization in T cells is the complex of cell surface molecules that assemble with the T cell receptor (TCR). The TCR complex typically contains multiple TCR polypeptides (α/β or γ/δ) in a complex with CD3 chains (ε, γ, δ, and ζ). Fluorescence microscopy has shown that the TCR is rapidly clustered at contact sites displaying peptide-MHC protein (pMHC) binding sites49; this phenomenon is most dramatically seen in the ultimate formation of supramolecular activation complexes (SMACs).50 Recent work has shown that the TCR complex can include multiple TCR heterodimers (α/β) and, interestingly, these complexes can exist in a range of clustered states.51 Clustering may be a mechanism for controlling the avidity and sensitivity of the TCR complex for its cognate antigen, and cluster formation may depend on the lipid environment in which the complex resides. Furthermore, clustered TCR complexes may serve not only as a mechanism for signal attenuation but also as the initial site of signaling—whereas the subsequently formed SMACs appear to be sites for terminating the signal as the TCR is degraded.52
An important component of the TCR counter-receptor, the MHC protein, also experiences lateral reorganization in the membrane. Both FPR26, 28, 53 and SPT30, 54, 55 studies have shown that the MHC protein is a highly mobile cell surface receptor, and its mobility is in part determined by its cytoplasmic tail. FRET56-58 and single-molecule fluorescence imaging59 studies have shown that MHC proteins also reorganize into oligomeric clusters. The lateral mobility of the lymphocyte receptor CD2 has been examined by FPR and SPT.41, 60, 61 Using both confocal FM and SPT, Douglass and Vale observed that CD2 associates specifically with membrane microdomains.22 The receptor traverses these domains with reduced lateral mobility, and other signaling molecules are found within these domains along with CD2. Other receptors, such as L-selectin62 and LFA-1,63 are also known to reorganize into clusters. The binding of multivalent ligands may mimic adhesion systems by enforcing surface receptor clustering.64, 65 In some cases, the functional consequences of this clustering may include activation and proteolytic shedding of the receptor.66, 67
RECRUITMENT
Like changes in the lateral organization of a membrane receptor, changes in the subcellular localization of the receptor can be critical for function.
Recruitment: Accumulation of a receptor in a specific subcellular region by passive or active mechanisms, usually involving interaction of the receptor with extracellular or cytoplasmic binding partners. (Figure 2)
2. Recruitment of membrane components. A: Cell surface receptors can be recruited to specific subcellular locations by interaction with extracellular ligands, including both cellular counter-receptors and binding sites in the extracellular matrix. B: Intracellular components, such as cytoskeleton-associated proteins, can also function to localize cell surface receptors.
The selectin family of cell surface receptors provides an excellent example of the importance of subcellular localization for receptor function. Selectins are found on a variety of leukocytes and are critical for the process of inflammation. The extracellular domain of the receptor contains a lectin binding site that is essential for receptor function. It has become clear, however, that the localization of the receptor on the cell surface is also critical for function.68 For example, L-selectin localizes to the microvilli of leukocytes, and experiments using selectin chimeras that contain the L-selectin binding domain and the transmembrane and intracellular domains of CD44—a cell surface molecule not generally found on microvilli—show that the localization of L-selectin to microvilli depends on the intracellular domain of the protein.69 Furthermore, the localization of L-selectin to microvilli depends on molecular interaction of the selectin intracellular domain with α-actinin, a cytoskeleton-associated protein.70 This molecular association is functionally important, as perturbations in the subcellular localization of L-selectin affect the ability of this receptor to mediate adhesion.70, 71 Other leukocyte receptors, such as the MHC protein on antigen presenting cells, are known to be associated with lipid raft domains, and the domains themselves appear to be actively recruited to the site of adhesion.72 In a similar fashion, both the SMACs and individual molecular components within the immune synapse—such as LFA-1, CD2, and the TCR—are directed via extracellular contacts and cytoskeletal mechanisms to the appropriate subcellular location, where an adhesive contact is formed.32, 50
DISPERSION
As described in the examples above, the recruitment of cell surface receptors to a specific subcellular location can be important for function. The converse of this process, i.e., molecular dispersion, can also be important for receptor function.
Dispersion: Conversion to (or maintenance of) a receptor in a randomly distributed state on the cell surface, generally by passive diffusion mechanisms. (Figure 3)
3. Dispersion of cell surface components. A: Ligands or receptors that are not attached or associated with localized components, such as cytoskeletal proteins, tend to become evenly distributed on the cell surface. B: The pool of dispersed receptors may be controlled by reversible association with cytoskeletal contacts or by clustering.
In the case of lymphocyte73 and related integrins,74 it appears that the resting state of the receptor has low lateral mobility. Both cellular stimulation and cytoskeletal disruption lead to a large increase in integrin lateral mobility and, thus, to dispersion of the receptor in the plasma membrane. One potential rationale for the effect of cellular stimulation on integrin mobility would be to “prime” the cell for adhesion by ensuring that the receptor binding sites are well distributed so as to foster binding. The lymphocyte receptor CD2 is also randomly distributed (i.e., disperse) on the surface of resting cells.38 Dispersion of adhesion receptors is often followed temporally by a localized recruitment and/or anchoring mechanism. For example, LFA-1 is ultimately recruited to the immune synapse as part of a SMAC50 and CD2 is selectively immobilized at sites of receptor ligation.41 One can therefore think of the dispersion mechanism as the counterpoint to receptor recruitment and anchoring.
ANCHORING
Altering the lateral mobility of a cell surface receptor can allow the molecule to reach its site of action—such as a contact or binding site. Once the receptor is at the proper location, its lateral mobility may be further modified to enhance receptor function. Some models propose passive mechanisms for supramolecular organization,75 but it is often found that adhesion proteins are immobilized by active mechanisms at the site of contact.33 Receptor immobilization can be accomplished by attachment to structural components of the cell, e.g., the cytoskeleton, effectively anchoring the molecule in place. Receptors with diffusion coefficients less than 0.1–1 × 10−11 cm2 s−1 may be considered as essentially immobile on the time scale of the cell, since such receptors would require many hours or days to explore the surface area of even a relatively small cell (vide supra). Therefore, receptor anchoring is an effective mechanism for positive reinforcement of molecular and cellular adhesive interactions.
Anchoring: Immobilization of a receptor at a specific subcellular location, generally through a change in molecular association of the receptor with cytoplasmic and cytoskeletal proteins. (Figure 4)
4. Receptor anchoring. A: Freely diffusing membrane receptors can become immobilized through association with the cytoskeleton. B: Association with the cytoskeleton can be direct, through binding of the receptor to a cytoskeletal component (primary), or indirect, through a linking protein that binds both a cytoskeletal component and the cytoplasmic tail of the receptor (secondary).
Adhesion receptor anchoring is an established mechanism of mobility regulation for several members of the integrin family. These heterodimeric adhesion receptors bind to a variety of counter-receptors, providing a physical link between the cellular cytoskeleton and the external environment. Measurements of integrin lateral mobility illustrate the molecular regulation of this process, and it is striking how many regulatory elements are conserved across different cell types and integrin subtypes. In fibroblasts, antibody-labeled β1 integrins diffuse rapidly, but the same integrins labeled with a counter-receptor, fibronectin, show a dramatic reduction in lateral mobility.74 Notably, the effect of ligation on integrin diffusion is observed only in integrins with intact cytoplasmic domains. The lateral mobility of the lymphocyte integrin, LFA-1 (αLβ2), has been studied using FM and SPT. The receptor is immobilized by cytoskeletal contacts in resting cells, and this anchored state can be disrupted by cell activation76 or cystoskeletal disruption.73 Several cytoskeleton-associated proteins appear to be involved in LFA-1 mobility regulation, including dynamitin77 and talin.78
Similar to the immobilizing effect of fibronectin ligation on β1 integrin diffusion,74 the lateral mobility of the β2 integrin LFA-1 is affected by binding to its counter-receptor, ICAM-1. Peters et al. observed that LFA-1 lateral diffusion is reduced when the receptor is labeled with ICAM-1, and hypothesized that the anchoring of bound LFA-1 would be expected to reinforce integrin adhesion.79 This observation was confirmed and extended by examining not only the diffusion of LFA-1 bound to ICAM-1, but also the mobility of different conformational epitopes of the receptor.15 Cairo et al. found that LFA-1 is indeed restricted in its lateral mobility on resting cells, but that activated conformations of the receptor are more mobile. These data fit a mass-transport model of LFA-1 diffusion, which may also be relevant for other adhesion receptor systems.48 A key observation from this study was the co-existence of multiple populations of receptors with different lateral mobilities. This heterogeneous diffusion profile suggests that the αLβ2 integrin exists in a dynamic equilibrium between its cytoskeleton-associated and freely diffusing forms.
We suggest that the observation of multiple states of lateral mobility coexisting at the cell surface is not unique to this system, and could well be a ubiquitous feature of membrane adhesion receptors. The most direct statement of this model can be found in Cairo et al. for LFA-1,15 although SPT experiments on other integrins have also found evidence for multiple populations of molecules with different diffusion coefficients (personal communication from M. Cho and H.-F. Chen).80 Similar observations of heterogeneous diffusion have been made for the lymphocyte receptor CD2.22, 41 It is important to recognize that this hypothesis is directly supported by even the earliest studies of membrane protein lateral mobility, in which FPR data on live cells typically showed a mobile and an immobile fraction—indeed, based on this evidence alone, we would have predicted that SPT experiments would reveal a heterogeneous population for most, if not all, adhesion receptors.
INTEGRATION OF MECHANISMS
We have seen several individual examples of mechanisms that affect lateral mobility, but few of these mechanisms could be said to operate independently. Indeed, it is likely that many adhesion systems integrate multiple mechanisms of mobility regulation to accomplish their function. Several key examples from T cell biology demonstrate this interplay: the integrin LFA-1, L-selectin, and the TCR complex. Each of these systems has been studied to the extent that the true complexity of its regulation has begun to be revealed.
Using a variety of methods, studies of the lymphocyte integrin LFA-1 have elucidated a complex network of elements coordinating T cell adhesion during lymphocyte rolling, extravasation, and antigen presentation. Some models of LFA-1 function have been controversial, most likely because the receptor's behavior is complex and highly regulated. It has been established that conformational changes in integrin heterodimers alter their affinity for ligand and that this alteration in affinity is necessary for the function of many integrin receptors, including LFA-1.81, 82 A longstanding debate has focused on whether conformational changes or receptor clustering events are more central to integrin function.34, 83, 84 As discussed earlier, because lateral mobility is required for the formation of receptor clusters, considerations of integrin lateral mobility regulation are necessary to resolve the issue.
Single-molecule studies of the relationship between LFA-1 conformation and lateral mobility have provided an emerging model that orchestrates these two factors into a single mass transport model.15, 48 Current data suggest that an intermediate-affinity receptor conformation is anchored, that cellular activation causes dispersion of the receptor, and that receptor ligation induces recruitment and anchoring of a high-affinity receptor conformation at the contact site in order to strengthen the adhesion.73 This model provides for negative regulation by the first anchoring mechanism and for positive regulation by dispersion and by the second anchoring mechanism (most likely, involving alternative cytoskeletal regulators).78 Reorganization of LFA-1 into multivalent complexes is also likely to be key to receptor function during these steps.63 Importantly, models with this level of resolution begin to approach a quantitative understanding of the system. By separately considering each conformational state of the receptor and the molecular interactions that affect its lateral mobility, we can view the overall system as a set of dynamic equilibria.15 This model is satisfying in its simplicity, and suggests that the cell can efficiently regulate integrin-mediated adhesion through modification of the binding affinity of the integrin to the relevant cytoplasmic proteins.82
L-selectin represents a second intensively studied adhesion system that integrates multiple mechanisms of mobility regulation. L-selectin-mediated adhesion is regulated by both receptor recruitment and receptor anchoring. Several lines of investigation show that recruitment of L-selectin, resulting in its localization to the tips of microvilli, is essential for the initiation of contact between the leukocyte and the vascular endothelium.69, 71 In this system, recruitment appears to be mediated primarily by the transmembrane and intracellular domains of the receptor. In addition to recruitment, anchoring of the receptor to the cytoskeleton appears to be required for leukocyte rolling.70 Anchoring is mediated by the interaction of the cytoplasmic domain of L-selectin with α-actinin.
The group of proteins that form the TCR complex is perhaps the most complex cell surface receptor on the T cell. The TCR participates in the formation of an immune signaling complex that includes many other receptors and cytoplasmic components, ultimately forming microclusters and supramolecular activation complexes (SMACs) (Figure 5).50 These processes require exquisite regulation of TCR lateral mobility, depending on receptor reorganization, recruitment, and anchoring mechanisms. The TCR has been found to exist in a mixture of clustered states on the resting cell, and can be reorganized to form multiple complexes with different stoichiometries with other surface molecules.51 This reorganization is thought to assist in controlling the fidelity of TCR response as the functional affinity of the cluster is altered.85 TCR signaling also leads to the recruitment of membrane protein and lipid components as well as a bevy of cytoplasmic signaling molecules.52 Finally, the recruitment of TCR complexes to the appropriate subcellular locations is an active process mediated by anchoring to components of the cytoskeleton.86
Integration of Mechanisms: Individual mechanisms of lateral mobility regulation integrate to organize complex structures, such as a SMAC, that forms during T cell-APC contact. A: Antigen presenting cell (APC) with dispersed MHC molecules and ligands such as ICAM-1. These cell surface components become organized when the APC encounters a T cell. B: T cell components, such as the TCR complex and LFA-1, may also be relatively dispersed on the surface of resting cells. C: Upon formation of a mature SMAC, MHC and ICAM-1 are recruited to the cell-cell interface through binding to their counter-receptors. On the surface of the T cell, LFA-1 is recruited to the site of interaction with ICAM-1 and then anchored by cytoskeletal contacts, and the TCR complex may be reorganized through recruitment to other co-receptors or lipid raft components.
CONCLUSIONS
The lateral mobility of cell surface receptors is an important regulator of cellular adhesion. We propose a general framework for understanding the role of lateral mobility in adhesion regulation. Our framework includes four interrelated mechanisms of regulation: reorganization, recruitment, dispersion, and anchoring. This framework allows for comparisons among different molecular adhesion systems and provides a useful classification of existing data. As more complete models of membrane receptor function and protein-protein interactions are elucidated, the measurement of lateral mobility will become increasingly important for understanding cellular adhesion mechanisms.
Acknowledgements
The authors would like to dedicate this article to the memory of Dr. Elkan Blout and to the memory of Dr. William Veatch. CWC acknowledges support from NSERC and the University of Alberta Faculty of Science. DEG acknowledges support from the NIH (HL32854 and HL70819).
REFERENCES
1Dustin, M. L.;Bromley, S. K.;Davis, M. M.;Zhu, C.Annu Rev Cell Dev Biol2001,
17,
133–157.
50Bromley, S. K.;Burack, W. R.;Johnson, K. G.;Somersalo, K.;Sims, T. N.;Sumen, C.;Davis, M. M.;Shaw, A. S.;Allen, P. M.;Dustin, M. L.Annu Rev Immunol2001,
19,
375–396.
65Gestwicki, J. E.;Cairo, C. W.;Borrok, M. J.;Kiessling, L. L.In Recognition of Carbohydrates in Biological Systems Part A:
General Procedures;
Y. C. Lee; R. T. Lee, Eds.;
Academic Press:
San Diego, CA,
2003, pp
301–312.
Please check your email for instructions on resetting your password.
If you do not receive an email within 10 minutes, your email address may not be registered,
and you may need to create a new Wiley Online Library account.
Request Username
Can't sign in? Forgot your username?
Enter your email address below and we will send you your username
If the address matches an existing account you will receive an email with instructions to retrieve your username
The full text of this article hosted at iucr.org is unavailable due to technical difficulties.


 ()
() ()
() ()
()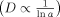 ,2 although more recent treatments suggest a stronger dependence
,2 although more recent treatments suggest a stronger dependence 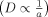 .18 From both models, we would predict that large receptor clusters would exhibit reduced lateral mobility. The formation of small clusters (e.g., dimers or trimers) might be difficult to discern in a live cell, however. In general, there are not many examples of large changes in receptor mobility due to clustering alone,42 and model studies of receptor aggregation using single-particle tracking have shown only small changes in lateral mobility.43
.18 From both models, we would predict that large receptor clusters would exhibit reduced lateral mobility. The formation of small clusters (e.g., dimers or trimers) might be difficult to discern in a live cell, however. In general, there are not many examples of large changes in receptor mobility due to clustering alone,42 and model studies of receptor aggregation using single-particle tracking have shown only small changes in lateral mobility.43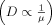 .2, 18 The situation is complicated, however, by the potential for heterogeneity within the membrane in the form of lipid rafts6 or other microdomains.16, 45 Work in model systems has clearly established the inherent preference of certain lipids to associate with one another,46 although direct observation of this phenomenon in vivo has been challenging.6 Membrane components diffusing through such microdomains could exhibit altered lateral mobility if the viscosity of the domain is significantly different from that of the surrounding lipid. Observations suggestive of this phenomenon have been found using single-molecule methods in live cells.22
.2, 18 The situation is complicated, however, by the potential for heterogeneity within the membrane in the form of lipid rafts6 or other microdomains.16, 45 Work in model systems has clearly established the inherent preference of certain lipids to associate with one another,46 although direct observation of this phenomenon in vivo has been challenging.6 Membrane components diffusing through such microdomains could exhibit altered lateral mobility if the viscosity of the domain is significantly different from that of the surrounding lipid. Observations suggestive of this phenomenon have been found using single-molecule methods in live cells.22

